物种形成是新物种从旧物种中分化出来的过程,即从一个种内产生出另一个新种的过程,相信很多人都想知道物种形成有什么条件,以下是由爱华网小编整理关于物种形成的条件的内容,希望大家喜欢!
物种形成的条件现代生物学关于种形成的研究对象大都是进行有性生殖的动植物,因而种形成的研究多集中于“生殖隔离”的起源问题上,即研究在一个种的群体内如何产生或分化出与原群体生殖上隔离的亚群体,后者就是一个新种的开始。
C·R·达尔文早就注意到地理因素在种分歧和种形成中的作用。P·M·哈蒙德1981年将与种形成有关的地理因素区分为13种情况,并归纳为4种模式:分布区重叠模式、分布不重叠模式、分布区相邻模式、分布区不重叠—相邻模式。
所谓“分布区重叠”是指形成不同种的原群体在地理分布上是连续的,所谓“分布不重叠”是指形成不同的种的原群体由于某种地理隔离因素而被分隔,所谓“分布区相邻”是指形成不同种的原群体之间有不完全的地理隔离。区分种形成方式主要依据群体初始分化和生殖隔离完成过程中的地理分布情况,至于生殖隔离完成(种形成过程结束)以后地理分布情况则并不重要。一般说来可以区分为三种不同的种形成方式:①分布区不重叠的种形成,②分布区重叠的种形成,③分布区相邻的种形成。
此外,根据种形成过程的特点还可区分出两种不同的种形成方式:①渐变的种形成,②量子种形成。分布区不重叠和相邻的种形成过程一般是渐变的,分布区重叠的种形成过程往往是“跳跃”的,即“量子种形成”方式。
分布区不重叠的种形成
一个广布的种,由于地理隔离因素而分隔为两个或若干个相互隔离的群体,由于这些被隔离的群体之间的基因交流大大减弱或
甚至完全停止,从而使各隔离群体之间的遗传差异逐渐增加,通过若干中间阶段(如形成族、亚种等)而最后达到生殖隔离。这样,原先隔离的群体就形成不同的种。由于在初始分化和生殖隔离完成阶段各个原群体的分布区不重叠,故名不重叠种形成。一旦生殖隔离完成,新种分布区即使再重叠(例如由于地理隔离因素消失),也不能交配或交配不育了,暗绿柳莺便是一个很好的例子。此鸟分布很广,可分五个亚种,形成为一个分布圈:①新疆亚种(P.t.viridanus),分布在中国新疆西部和北部,国外伸展到中亚部分地区、西伯利亚西部直到欧洲北部;②拉达亚种(P.t.ludloui),分布在克什米尔地区,其北端与新疆亚种衔接,两者相互杂交;③指名亚种(P.t.trochi-loides),分布在中国西藏南部喜马拉雅山地区,云南西部和四川南部。其西端与拉达亚种衔接杂交;④甘青亚种(P.t.obscuratus),分布在中国甘肃、青海和四川西部,在四川和指名亚种相遇杂交;⑤东北亚种(P.t.plumbei-tarsis),分布于中国东北、内蒙古和西伯利亚东部,其南端与甘青亚种间未发现有衔接区,其西部与新疆亚种在萨彦岭一带相遇,则不相杂交,已形成生殖隔离(图1)。
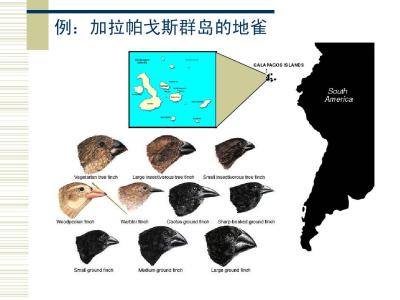
不重叠种形成过程通常是长时间的,不可能自始至终直接观察到。这种物种形成方式的提出,多半是根据间接的证据,例如不完全种(族、亚种或半种)的存在,某些生物种的地理分布等。一些大洋中的群岛对于某些陆地生物来说是不重叠种形成的合适环境。最典型的例子是达尔文曾考察过的南美西岸的加拉帕戈斯群岛上的种类丰富而又歧异的动植物区系。例如被称为达尔文莺的小鸟有10~14种之多,达尔文认为这些不同种的莺来自一个从大陆迁徙到该群岛的祖先种。由于群岛与大陆隔离和群岛之间的隔离使得迁入的鸟群分异,并形成不同的种,加拉帕戈斯群岛上有3种龟,分别栖居于被海洋分隔的3个小岛上,这也是地理隔离因素所促成的不重叠种形成过程的间接证据。
在海洋生物中也有许多由于地理隔离而形成不同亚种的实例。某些物种在北太平洋东西两侧有不连续的分布,例如某些软体动物和鱼类在北美西海岸是一个亚种而在苏联远东地区和日本北部沿海是同种的另一个亚种,原因是从白令海峡流过的来自北极的寒冷海流,构成了海洋生物的分布障碍。某些海洋动物在北大西洋和北太平洋对称地分布而在北极中断。例如鳕鱼分布于北大西洋,在北太平洋则有同种的另一亚种,但在北极却没有鳕鱼。显然北极的寒冷是分布中断的原因,但可能在某一地质时期北极地区曾经是比较温暖的,两大洋北部的生物曾有连续的分布。中美的某些生物,由于巴拿马地峡的阻隔,而沿着东西两岸分布着两两对称的姐妹种。所谓姐妹种就是形态上非常相似而生殖上隔离的完全种。在地峡形成以前它们的分布可能是连续的。
分布区相邻的种形成
分布于渐变的环境或不均一的环境中的广布种,在分布区的不同部分形成相互接触而又有一定程度的隔离(不完全的隔离)的群
体。群体间在接触区域有一定程度的基因交流,但群体的分布区的中心部分之间的基因交流很弱或甚至完全没有。这样,群体之间的遗传差异会逐渐增大,由此而形成不同的种。某些间接的证据可以说明相邻的种形成方式的存在。例如由呈环链状分布的若干亚种组成的所谓“环布种”,相邻的亚种之间有部分的基因交流,环链两端的亚种之间可能分布重叠,但往往不能互相杂交或杂交不育(生殖隔离)。一个典型的实例就是环绕北极的鸥,它是由若干亚种组成的,更新世以后分布于北太平洋的亚种越过北美和北大西洋而与西欧的亚种相遇,但仍保持显著的差异而不婚配(图2)。
由若干相邻分布的亚种组成的广布种,各相邻的亚种之间有一定的基因交流,但分布两端的亚种(1与5)通过长期的遗传差异的积累而可能达到生殖隔离(即使分布区重叠也不能杂交)。
分布区重叠的种形成
一些证据表明,生殖隔离可以由于生物种的行为差异、发情或开花季节差异、寄主,以及其他原因而导致在那些分布区并非完全不重叠的群体内产生。有些学者提出一些证据证明植物的某些新种的形成是没有地理隔离的情况下达到生殖隔离的。
但分布区是否重叠往往很难区分。例如人体寄生有3种虱:阴虱寄生于腋毛和阴毛中,人虱分布于躯体(主要在衣服上),头虱生于头发上。若从整个人体看,3种虱的分布区是重叠的,但若从人体的局部部位看,它们又是不重叠的。分布区是否重叠一般应根据具体的生物对象的活动能力和行为等特点来判断。
分布区不重叠的和相邻的种形成方式
新种产生于遗传上连续的单个群体之内(分布区重叠的种形成)或者产生于有基因交流的相邻分布的一系列群体之中(分布区相邻的种形成)。在没有任何阻止基因交流的外在的隔离因素的情况下,新种从母群体中分化产生出来的一些证据多是从观察研究“成对种”和“初始种”的分布和区分特征而获得的。“成对种”和“初始种”被认为是新形成的或形成中的物种,它们常是研究物种形成的对象。但是根据它们的地理分布和区分特征来推断种形成方式还存在许多问题。例如“成对种”或“初始种”的现有的分布区相邻的状况也可能是种间竞争所造成的地理分布上的相互排斥,如是,则其相邻的分布乃是种形成的结果或种形成后的现象。母种与新种之间的区分特征中哪些是种形成过程所涉及的,哪些是种形成以后产生的,也很难判断。
根据学者们提供的资料,可将分布区重叠的和相邻的种形成归纳为两种可能的方式:
①两阶段的种形成过程。在一个遗传上连续的群体之内首先发生种内分异,经过相当长时间的种内分异的积累而最终在群体内产生生殖隔离的新种。
②一步“跳跃”的过程。新种从母群体中突然产生,即通过染色体突变或基因突变或其它机制在较短时间内产生与母群基因交流阻断的新类型或新种,一举而达到生殖隔离。
关于第一种方式可以举出下面一些事实。分布区重叠的种形成的实例如某些寄主专一的昆虫寄主族是近期形成的半种,是种内分异的结果。
1975年,G·L·布什在研究果蝇的寄主族(指同一物种的有不同的寄主专一性的族群)后指出:在同一物种内寄主的转换可以造成分布区重叠的寄主族。布什认为,对于果蝇来说寄主转换只需要很少的基因突变,而寄主的改变只是种内分异的第一步,其后是生态小生境的改变并伴随寄主族之间、新寄主族与母群体之间的生殖的障碍。例如,果蝇的交配只发生在其寄主的果实上,不同的寄主族之间发生交配的可能性很小,这样,寄主族之间的分异就会和地理隔离造成的效果一样。
对澳洲蝗虫的研究提供了分布区相邻的种形成方式的例证。1978年,M·J·D·怀特研究了澳洲蝗虫的相邻分布的近缘种和染色体族之后提出了“原地静态”的种形成方式,他认为,染色体重排(染色体融合、分离、倒位、易位、断裂、缺失等)在种内初始分异中起重要作用。澳洲蝗虫是失去飞翔能力的、种内核型变异很大的昆虫,大约有200种之多。在一些种(例如在 Vandiemenella viatica)之内有许多核型不同的、地理上相邻分布的染色体族。有一些核型不同的近缘种的分布区也是相邻的。这些相邻分布的族或种的分布区往往有明确的边界,互不重叠。研究还发现染色体族之间存在着程度不同的生殖障碍。例如,各族之间的杂合子不育或生育力低,从而在各染色体族之间形成一狭窄的“杂交带”。由于基因交流的阻隔,染色体族有可能发展为生殖隔离的新种。
但对于这些分布区相邻的染色体族和种的形成的解释还有一些不同的看法。首先是关于染色体重排在种形成过程中究竟起多大作用还难以评价。在澳洲蝗虫中并非所有的近缘种在核型上都有区别。另一个疑问是:相邻的分布是否可能为种间竞争排斥的结果,即种形成后的现象。还有一个问题是:种内分异所造成的族(半种)之间的生殖障碍(不完全的生殖隔离)如何发展为完全的生殖隔离。达尔文和A.R.华莱士都曾提出过生殖障碍可以通过自然选择而逐步发展的假说。如果种内分异产生出多种表型,如果中间类型(杂种)的适应值相对较低,那么通过“中间类型”或“杂种”的淘汰绝灭而由生殖障碍发展为完全的生殖隔离。
第二种跳跃的种形成方式是不经过种内分异而一步达到生殖隔离的过程。例如在植物中,多倍体的种可能是从其二倍体的亲种中快速产生的。H.刘易斯(1966年)认为木本植物的染色体基数在科或亚科级的大分类单元中是恒定的,在草本植物中染色体基数在属一级的类群中是恒定的。植物的染色体基数的改变可能涉及高级分类群的形成,因而是跳跃的。
物种形成的域种化异域种化:同一物种由于地理隔离,分别演化为不同的物种。
同域种化:同一物种在相同的环境,由于行为改变或基因突变等原因而演化为不同的物种。
边域种化:种化过程中,一个小族群由于某种原因和原来的大族群隔离;隔离时,小族群的基因经历剧烈变化;当小族群再跟大族群相遇时,已经形成不同物种。
临域种化:两个种化中的族群虽然分开,但是相邻;从一极端到另一极端之间的各族群都有些许不同,但彼此相邻的两族群之间仍能互相杂交;不过,在两边最极端的族群已经差异太大而形成不同的种类。
物种量子形成思想20世纪30年代,群体遗传学家S·赖特1932年所提出的“适应峰”概念就包含有量子种形成的思想。赖特用类似地形图的“适应图”(图3)来形象地表示生物
种的基因组合对环境的适应状态。赖特认为自然界中的种各自占据着适应图中的“峰”(标有“+”号的部位),这些“峰”代表生物种的基因组合及其表型特征具有最大适应值,而处于“谷”(标有“—”号的部位)中的基因组合适应值最低。由“峰”到“谷”之间的等高线代表适应值由高到低逐步下降。新种的形成意味着从一个“峰”越过“谷”而达到另一个“峰”,这只有通过显著的进化改变或快速的“跳跃”才能实现,因为缓慢的过渡要越过适应“谷”(适应值最低的状态)是不可能的。
古生物学家G·G·辛普森(1944年)鉴于化石记录中某些高级分类单元在较短的地质时间内出现而提出“量子进化”的概念。他认为生物种通过跳跃式的“量子进化”就能越过适应阈限,进入新的适应带,从而发生适应辐射和产生高级分类单元。但辛普森认为“量子进化”可以通过分枝的种形成,也可以通过线系进化而实现。
某些生物学家(例如G·G·辛普森、E·迈尔等)认为,除了渐变的种形成方式以外,还可能存在快速的、跳跃式的种形成方式。另一些生物学家(如斯坦利)则认为种形成不可能是渐变的过程,只能是“跳跃”式的、“相对快速”的过程。处于“相对稳定”状态的种,通过“突然”的改变而分化出新种的过程称为量子形成。
1963年,这个词最先由V·格兰特提出,S·M·斯坦利把它定义为“大部分进化集中发生在种形成的初期的相当短的时间(相对于种的寿命而言)之内的种形成过程”。关于种形成的方式是渐变论者与间断平衡论者之间争论的焦点。渐变论者认为渐变的种形成是一般的、普通的,快速、跳跃式的种形成是极特殊的情况;间断平衡论者则认为种的形成只能是“跳跃”式的。
目前,关于量子种形成问题的争论焦点在于快速种形成的具体机制。一些学者强调这种机制是“大突变”,甚至认为一个或少数几个突变就能产生一个新种或新的高级分类单元。这实际上又回到了20世纪初期德·弗里斯的“突变论”。
有的学者则提出另一种快速种形成的可能机制:在分隔状态下的小群体,随机因素有助于快速进化和种形成。如果一个广布的种由许多小群体组成,小群体之间有不完全的隔离,则随机因素会造成小群体对总体的大的偏离和小群体之间的遗传差异。但由于小群体的不稳定也增大了绝灭的可能性。
看过“物种如何形成”的人还看了: