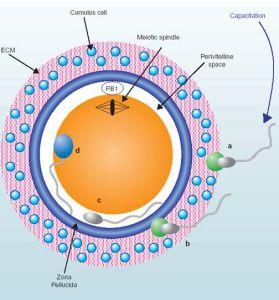
受精,指卵子和精子融合为一个合子(受精卵)过程,是有性生殖的基本特征,普遍存在于动植物界。在细胞水平上,受精过程包括卵子激活、调整和两性原核融合3个主要阶段。激活可视为个体发育的起点,调整发生在激活之后,是确保受精卵正常分裂所必需的卵内的先行变化;两性原核融合起保证双亲遗传的作用,并恢复双倍体。在分子水平上,受精不仅启动DNA的复制,而且激活卵内的mRNA、rRNA等遗传信息,合成出胚胎发育所需要的蛋白质。
受精_受精 -概述
受精的早期事件
受精(fertilization):受精(fertilization)是卵子和精子融合为一个合子的过程。它是有性生殖的基本特征,普遍存在于动植物界,但人们通常提到最多的是指的动物。动物受精在细胞水平上,受精过程包括卵子激活、调整和两性原核融合3个主要阶段。激活可视为个体发育的起点,主要表现为卵质膜通透性的改变,皮质颗粒外排,受精膜形成等;调整发生在激活之后,是确保受精卵正常分裂所必需的卵内的先行变化;两性原核融合起保证双亲遗传的作用,并恢复双倍体,受精不仅启动DNA的复制,而且激活卵内的mRNA、rRNA等遗传信息,合成出胚胎发育所需要的蛋白质。
受精包括精子的获能(capacitation)、顶体反应(acrosomereaction)、皮层反应(corticalreaction)、原核形成和融合等过程。
通常一个卵子和一个精子结合,多精进入会形成多余的分裂极和纺锤体,导致细胞异常分裂而使胚胎发育终止。受精后通过两种机制阻止多精进入,一是膜瞬间去极化;二是通过皮层反应,破坏精子受体和形成受精膜。受精后精子的核、中心粒(鞭毛基粒)和线粒体均注入卵中。线粒体之所以表现为母系遗传,是因为受精卵中只有仅母本线粒体可以存活。
许多昆虫、软体动物、软骨鱼、有尾两栖类、爬行类和鸟类在受精时有多于一个精子进入卵内,但只有一个雄原核与雌原核发生融合,参与发育,其余的精核逐渐退化消失。有时多余的精于也发生一些畸形的分裂,但不妨碍正常卵裂和发育。这种多精受精称为生理性多精受精。
受精_受精 -动物的受精
动物受精
受精是卵子和精子融合为一个合子的过程。它是有性生殖的基本特征,普遍存在于动植物界。
在细胞水平上,受精过程包括卵子激活、调整和两性原核融合3个主要阶段。激活可视为个体发育的起点,主要表现为卵质膜通透性的改变,皮质颗粒外排,受精膜形成等;调整发生在激活之后,是确保受精卵正常分裂所必需的卵内的先行变化;两性原核融合起保证双亲遗传的作用,并恢复双倍体。在分子水平上,受精不仅启动DNA的复制,而且激活卵内的mRNA、rRNA等遗传信息,合成出胚胎发育所需要的蛋白质。
研究简史
1875年德国生物学家O.赫特维希首先在海胆上发现从精子入卵至雌雄两原核融合的受精过程,胚胎学上争论200余年的唯卵和唯精学说,至此才得到合乎事实的解答。1883年比利时生物学家E.van贝内登发表二价马副蛔虫受精细胞学的研究论文,肯定了赫特维希的在遗传上父母贡献均等的理论,并使精、卵合作的研究更为深入。在马副蛔虫合子第一次分裂的纺锤体上,可看到四条染色体,其中两条来自父方,两条来自母方。因此,他认为染色体有定形、有定性、有定数和有系统,父母的染色体通过精卵的融合传给子代。后来,德国生物学家T.H.博韦里在马副蛔虫上的工作进一步巩固了上述理论,把染色体看作是遗传信息的载体。
20世纪以来,受精研究转向探讨两性配子结合的机制。美国学者F.R・利利根据沙蚕和海胆上的研究,首先指出卵子分泌出与接受精子有关的物质,他称之为受精素。40年代前后,另一美国学者A.泰勒就受精素的生物学、化学和免疫学特征展开了一系列工作,进一步强调卵子成熟过程中排出物对受精的重要意义。与此同时,德国学者M.哈特曼认为在海胆受精过程中,不但卵子能排出雌配素,精子也能排出雄配素,两者相互抗衡的程度决定着受精成功与否。不久,在两栖类上,发现卵外胶膜在受精中的作用。1956年,中国实验细胞学家朱洗等根据中华大蟾蜍的实验,提出输卵管分泌的卵外胶膜,为雌雄配子实现受精所必需。在哺乳动物方面,1951年张明觉和C.R.奥斯汀分别同时提出精子必须在雌体生殖道逗留一段时间,获得穿入卵子的能力――获能,才能有效地使卵子受精。精子获能的发现使人们找到过去哺乳类卵子离体受精不成功的原因,从而把高等哺乳动物和人类卵子受精的研究推向一个新阶段。
受精_受精 -人工授精
人工授精必须具备一定条件,其中最基本的条件是女方至少有一边的输卵管是完全通畅的(经输卵管碘油造影或者宫腹腔镜手术检查证实),女方有自发排卵或经促排卵治疗可以排卵;而男性的精子必须要有一定数量的活精子,只有这两个条件都具备,才有可能成功受孕。
受精方式
◆体内受精和体外受精
受精与卵裂
凡在雌、雄亲体交配时,精子从雄体传递到雌体的生殖道,逐渐抵达受精地点(如子宫或输卵管),在那里精卵相遇而融合的,称体内受精。凡精子和卵子同时排出体外,在雌体产孔附近或在水中受精的,称体外受精。前者多发生在高等动物如爬行类、鸟类、哺乳类、某些软体动物、昆虫以及某些鱼类和少数两栖类。后者是水生动物的普遍生殖方式,如某些鱼类和部分两栖类等。
自体受精和异体受精多数动物是雌、雄异体的,雌、雄两性是分开的。有些动物是雌、雄同体的,同一个体既能产生卵子,也能产生精子。在雌雄同体的动物中,有些是自体受精的,即同一个体的精子和卵子融合,如绦虫;有些仍是异体受精,即两个不同个体的精子和卵子相结合,如蚯蚓。
◆单精受精和多精受精
通常,只有一个精子进入卵内完成受精,称单精受精,如腔肠动物、棘皮动物、环节动物、硬骨鱼、无尾两栖类和哺乳类动物。这类卵子一旦与精子接触,就立即被激活并产生一系列相应的变化,阻止其他的精子入卵。如果因为卵子的成熟程度不适当等原因,而有一个以上的精子进入这类卵子,即所谓的病理性多精受精,则卵裂不正常,胚胎畸形发育,迟早必归天殇。有些卵子在正常受精情况下,可以有一个以上的精子进入卵子,但只有一个精子的雄性原核能与卵子的雌性原核结合,成为合子的细胞核,其余的精子逐渐退化消失,称生理性多精受精。如昆虫、软体动物、软骨鱼、有尾两栖类、爬行类和鸟类的受精。
受精过程
动物的精子不像低等植物如苔藓植物的精子有明显的趋化性,而是靠自身主动运动或依靠生殖道上皮细胞的纤毛运动抵达卵子附近。
精子获能和顶体反应
已知许多哺乳动物精子经过雌性生殖道或穿越卵丘时,包裹精子的外源蛋白质被清除,精子质膜的理化和生物学特性发生变化,使精子获能而参与受精过程。
哺乳动物的获能精子接触卵周的卵膜或透明带时,特异地与卵膜上的某种糖蛋白结合,激发精子产生顶体反应:顶体外围的部分质膜消失,顶体外膜内陷、囊泡化,顶体内含物包括一些水解酶外逸。顶体反应有助于精子进一步穿越卵膜。在海胆卵上,激起精子顶体反应的是卵周胶膜中的某种多糖物质。绝大部分卵的外周都有卵膜,各种卵膜厚度不一,主要组分是粘蛋白或粘多糖;只有少数是裸卵,如腔肠动物的卵。
精子穿越卵膜时,出现先粘着后结合的过程。前者为疏松附着,不受外界温度干扰,无种的专一性,粘着期间,顶体内膜上的原顶体蛋白转化为顶体蛋白,顶体蛋白有加速精子穿越卵膜的作用;后者是牢固的结合,能被低温干扰,具有种的专一性。在海胆精子质膜上已分离到一种能与卵膜糖蛋白专一结合的蛋白质,称作结合蛋白,分子量约30000。
卵子的激活
精子一旦与卵子接触,卵子本身也发生一系列的激活变化。在哺乳动物卵上,则表现为皮层反应,卵质膜反应和透明带反应,从而起到阻断多精受精和激发卵进一步发育的作用。皮层反应发生在精卵细胞融合之际,自融合点开始,皮质颗粒破裂,其内含物外排,由此波及整个卵子的皮层。卵质膜反应是卵质与皮质颗粒包膜的重组过程。透明带反应为皮质颗粒外排物与透明带一起形成受精膜的过程,卵膜与质膜分离,透明带中精子受体消失,透明带硬化。
精卵融合
精卵细胞融合时首先可以看到卵子表面的微绒毛包围精子,可能起定向作用;随即卵质膜与精子顶体后区的质膜融合。许多动物的精子头部进入卵子细胞质后即旋转180°,精子的中段与头部一起转动,以致中心粒朝向卵中央。接着雄性原核逐渐形成,与此同时中心粒四周产生星光,雄性原核连同星光一起迁向雌性原核。精子中段和尾部不久退化和被吸收。卵子细胞核在完成两次成熟分裂之后,形成雌性原核。雌、雄两原核相遇,或融合,即两核膜融合成一个;或联合,两核并列,核膜消失,仅染色体组合在一起,以建立合子染色体组,受精至此完成。
生化变化
卵受精之前,代谢水平很低,无DNA的合成活动,RNA和蛋白质的合成都极少。因此排出的卵子,如果未受精,很快就夭折。
当精子与卵子表面结合时,卵子的代谢速率迅速提高,并开始合成DNA。有关卵子激活的详细机理还不清楚,只知精子仅起到打开程序开关的作用。除了精子,一些其他非专一的化学的或物理的处理,也能使卵激活,例如针刺蛙卵,也能使之激动。激动的起始无需任何新蛋白质的合成。
在海胆卵激活的早期阶段,质膜对钠离子的通透性增加,钠离子大量内流,致使质膜在数秒钟之内去极化;钙离子自细胞库存中释放,使20~30秒钟之内卵内游离钙离子量迅速增加多达100倍;随着钠离子内流,氢离子外流,致使一分钟之内卵pH值明显增加。这些离子的变化,诱发皮层反应,导致阻断多余精子入卵,并激起卵的进一步发有。卵内游离钙离子的增加,赶到激活卵内钙调素的作用,由此进一步激活卵内其他的蛋白质。随即出现蛋白质合成量的增加,DNA也开始复制。
在海胆卵上,钙离子也可能是通过钙调素,激活卵质膜上的某些专一转运蛋白质,使氢离子向细胞外输出。卵内pH值的增加,会引起蛋白质合成速率增加和DNA的复制。这种氢离子的外流依赖于钠离子的内流。这些蛋白质的合成并不依赖于RNA的新合成,而是预存卵中mRNA的去掩盖以及核糖体激活的结果。
受精机制的研究,是人类有效掌握和控制有性生殖动物繁殖和育种的基本保证之一。人类“试管婴儿”的诞生只是少数成功的事例。由于受精和进一步正常发育机制方面尚存在许多悬而未决的问题,许多动物的体外受精的尝试均遭失败。这些失败表明基础理论研究的重要。
受精_受精 -植物的受精
受精是两种配子融合成为合子的过程。由合子发育成一具有双亲遗传性的新个体。受精是有性生殖的中心环节。
高等动植物的雄性和雌性亲本(即父本和母本)的遗传特性,是由具单倍染色体的精子和卵通过受精而传到子代的。由精子和卵融合产生的新个体,恢复了象亲代一样的二倍染色体的数目,继承了亲代双方的遗传性,同时,由于亲本双方遗传物质的重新组合,还有可能表现出新的性状。所以通过受精产生的子代,既有亲代遗传的特性,也表现有个体的特异性。因而,受精不仅在维持物种的延续上有重要的意义,而且也是生物进化的一个重要的因素。
基本方式
有三种:①同配生殖(同型配子配合);②异配生殖(异型配子配合);③卵式生殖(精卵配合)。
受精的三种形式反映了融合的两种配子从很少分化到有明显的分化的进化过程。
◆同配生殖
被子植物受精
融合的两种配子在形态和大小上是完全相似的,其中有两种情况:①同宗的个体(由一个个体经无性生殖或营养生殖所产生的后代)所产生的配子可以互相配合,叫做同宗配合;②甲宗的配子只能与乙宗的配合,叫做异宗配合。后者配子虽然大小、形态相同,但在生理上有差别。异宗配合的两种配子和产生它们的植物体通常分别用正号“+”和负号“-”表示。绿藻中衣藻属的某些种是异宗配合的。这些种的配子有?配子之间才能发生凝集和进一步的融合。真菌中的黑根霉以及毛霉属的一些种也可见到异宗配合生殖。当“+”系和“-”系菌丝相遇,在相对的位置可以产生原配子囊,而后相对的不同交配型的配子囊的原生质体融合。
◆异配生殖
融合的两种配子在大小上有固定的差别,这是性细胞进一步分化的表现。例如绿藻中的海松属,在一个重复分叉的个体上产生两种配子囊,在一种配子囊中产生具有二根鞭毛的形体较大的配子,另一种配子囊中产生与前者形态相似而形体小得多的配子,大的称为雌配子,小的称为雄配子。
◆卵式生殖
融合的两种配子在结构、能动性和大小上都有显著的差别。雄配子通常称之为精子,植物界许多类群的精子是具鞭毛和有运动能力的。精子的核大而细胞质少;高度分化的雌配子称为卵或卵球,卵细胞的细胞质极为丰富。从低等的藻类、真菌到种子植物都有卵式生殖,而同配生殖、异配生殖只发生在藻类和菌类。在同型配子配合和异型配子配合的受精方式中,性细胞都具有鞭毛,融合的两种配子可在水中游动相遇并实现受精。在卵式生殖中,受精的雌配子一般是不动细胞,性器官的结构也较复杂。受精的完成有多种不同的方式。团藻属的精卵配合是卵式生殖中最简单的代表。
在团藻的多细胞个体中可同时形成雄性和雌性两种配子。由一个雄性原始细胞经多次纵分裂产生许多细长而具两根鞭毛的精子;卵则是由一个雌性原始细胞增大而成,成熟时为球形,无鞭毛,游动的精子游至被胶质鞘包被的卵中,由一个精子与卵融合完成受精作用。
藻类和真菌植物还有其他精卵融合的方式。例如真菌中的水霉属,雌性器官――卵囊在菌丝的末端形成,在卵囊中发育出一至多个卵;产生精子的精子器在卵囊的附近产生,精子数量多,不能游动。在受精时,由精子器长出的受精管穿入卵囊并将精子射出达到卵旁而实现受精。绿藻中无隔藻属的性器官与水霉基本相似,但精子能动,成熟的卵囊中只有一个卵。精子从精子器释放出来以后,通过卵囊喙状开口进入卵囊与卵融合。
在苔藓和蕨类植物中,精子和卵都包藏在多细胞的性器官中,受精还不能离开水的条件。地钱的雌性器官形如瓶状,称为颈卵器。颈卵器与精子器分别生于叶状配子体的托上。地钱是雌雄异体的。在颈卵器中只产生一个卵,而在精子器中产生许多长形卷曲而具二根鞭毛的精子。当精子器成熟时,精子释放出来,如果植物体背面有水,精子可以游至颈卵器与卵实现受精。蕨类植物的颈卵器和精子器是在同一个配子体的下表面产生,能游动的精子经配子体表面的水层游进颈卵器内。
种子植物生殖器官的结构更为复杂,在有性生殖过程中产生了花粉管,借助花粉管,受精不再依赖水的条件,这是种子植物适应陆地生活的一个重要因素。产生雌配子的雌配子体在大孢子囊即胚珠的珠心处发育。裸子植物中的大多数仍保留颈卵器的结构,卵在颈卵器内形成。在小孢子囊中发育的花粉粒,即前期的雄配子体。在雄配子体中形成两个精子。精子多不具鞭毛,但在苏铁目和银杏目仍具游动的精子。被子植物的雌配子体称为胚囊,卵直接在其中产生;精子完全失去鞭毛。这些特点决定了种子植物的受精必先经过传粉。裸子植物的胚珠是裸露的,经过传粉,花粉被送到胚珠的珠孔处,而在被子植物中,胚珠着生在雌蕊的子房内,花粉粒不能直达胚珠而是落在柱头上,再由萌发生长的花粉管将精子输送到雌配子附近。在裸子植物中,进入颈卵器的两个精子只有一个与卵融合。但在被子植物中进入胚囊的两个精子则分别与卵和极核融合。这就是被子植物特有的双受精。
受精_受精 -受精过程中的识别作用
20世纪40年代已认识到衣藻的有性生殖是由一种称为配素的物质所控制。后来人工培养的试验进一步明确配子状态的细胞释放至培养基的物质能引起相对的交配型的配子凝集;即从“+”系的细胞释放的物质可使“-”系的细胞凝集,反之,从“-”细胞释放的物质可凝集“+”细胞,在同性凝集之后,并不发生细胞融合。这种物质称为同种凝集素,它是一种含糖蛋白的物质,存在于鞭毛的表面的微小泡中,它不仅有性的专一性,而且有种的专一性,无疑是涉及配子的粘着和识别反应的物质。
在真菌的毛霉目的有性生殖中存在类似的识别反应。例如毛霉属异宗配合时是通过“+”系和“-”系配子囊的融合,人工培养的实验已经证明,控制异宗配子囊的融合分两步。首先要由三孢酸诱导产生两种交配型的原配子囊,而三孢酸只有在两种交配型存在的情况下才能形成,因为一种交配型产生的前体只能被相对交配型转变为三孢酸,即“+”系产生的前体只能被“-”系转变为三孢酸,“-”系产生的另一前体只能被“+”系转变为三孢酸。然后不同交配型的原配子囊相遇才发生融合。这一识别反应,推测是由于在配子囊顶端区域的壁上存在一种含糖蛋白的识别物质。
苔藓和蕨类植物的卵包藏在颈卵器中,游动的精子到达颈卵器和卵结合必需游至一定距离,精子能“发现”卵,显然是存在一定的相互作用的物质。在这两大类群的植物中,精子的向化性已被肯定。向化性指精子趋向卵或颈卵器产生的化学物质。W.F.P.普菲费尔最早研究欧洲蕨(Pteridium aquilinum)的精子向化性的实验证明,由颈卵器产生的L-苹果酸是有效的吸引剂。它将含有自来水的(包含10 %琼脂) 10%L-苹果酸钠的滴管插入精子的悬液中,苹果酸盐离子立刻从滴管中有级差地扩散到四周的介质中,精子在一秒钟内便移向滴管的口,即向苹果酸盐来源的方向移动。在缺少苹果酸盐的自来水中,精子是随意行动的;如果在均质的苹果酸盐的溶液中,即在级差不存在时,精子在溶液中也保持均匀的分布。在问荆曾试验过12种有机酸,精子仅对L-苹果酸和酒石酸敏感。
在被子植物中,精子由花粉管传送至胚囊。对花粉管通过花柱定向生长的原因,曾有各种解释。有的推测也是向化性因素的控制,即认为钙是吸引花粉管生长的因素;有的认为花粉管早期生长受营养核中DNA转录的mRNA所控制;还有的假设花粉管定向生长决定于柱头至子房电位差等等。这是一个尚无定论的问题。
花粉与雌蕊的相互识别对被子植物实现受精起决定性的作用。因为选择雄配子的首先是雌蕊组织,而不是卵。
在被子植物中,柱头和花柱有“筛选”出自己最合适的花粉的作用。大多数植物广泛表现为同一种内的异花受精,如果遗传上差异较大(如属间或种间的关系)或者遗传上差异太小(如同一朵花、同一个体不同花之间的自交)的个体之间交配则不亲和,这种远缘杂交不亲和以及自交不亲和的现象,是被子植物在长期进化过程中发展起来的既有利于维持种的稳定性又能保持其生活力的一种适应特性。
落到柱头上的花粉是被“接受”或是被“拒绝”决定于它们之间的相互识别。如果是亲和的,植物的花粉粒便在柱头上萌发出花粉管长入柱头,并沿一定的途径通过花柱到胚囊,在那里释放出精子;如果植物的花粉与柱头是不亲和的,雌蕊将抑制花粉的萌发或花粉管的生长而阻碍受精。
植物受精的不亲和性是由两亲本的基因型所决定,并表现出生理上的抑制反应。自交不亲和性的生理基础根据对十字花科和菊科的一些植物的研究,认为是花粉与雌蕊组织间识别反应的结果。与识别有关的物质,在花粉的一方是存在于花粉壁的蛋白质;在雌蕊一方已确定的是覆盖在柱头表面的蛋白质薄膜。
花粉有外壁和内壁。组成外壁的物质是类胡萝卜素和类胡萝卜酯氧化多聚化而形成的孢粉素。在外壁物质所形成的结构之间有由绒毡层合成和运入的蛋白质,当内壁发育沉积果胶纤维素时,由花粉本身细胞质合成的蛋白质并入其中。这样,当花粉成熟时,其外壁有孢子体(绒毡层细胞)基因型控制产生的蛋白质;内壁有配子体(花粉中细胞)基因型控制产生的蛋白质,这些都是起识别作用的蛋白质。
关于雌蕊对花粉的识别方面,目前对干型柱头(柱头成熟时无液体分泌物)的认识比较清楚。例如在十字花科和菊科的植物,柱头的表面产生乳头状单细胞,它具果胶纤维素的壁及一层不连续的角质层。乳突状细胞在发育过程中,通过壁和角质层还分泌一些物质形成一层薄膜覆盖在表面,此薄膜主要成分是蛋白质,它不仅提供了一个有利于“捕捉”花粉的粘的表面,而且具有识别“感受器”的特点。当与花粉壁蛋白结合后接着发生“接受”或“拒绝”反应。
花粉落在柱头上,从柱头吸水膨大,在接触后的几分钟内,花粉壁蛋白质即释放出来与柱头表面的蛋白质薄膜结合,如果是亲和的结合,花粉萌发,由于酶的作用,柱头的角质层被溶解,花粉管进入柱头并在花柱中顺利地生长。在不亲和的情况下花粉管生长停滞,或不能进入柱头(孢子体系统控制),或花粉管在花柱中生长停顿和破裂(配子体系统控制)。萝卜、白菜、大波斯菊和向日葵等植物在不亲和的自交中,传粉几小时后,柱头的乳突状细胞中即产生胼胝质的凸透镜状结构;而在亲和的交配中则没有胼胝质反应。这种胼胝质反应可以作为在一定亲缘关系范围内检验亲和性的指标。一种花粉落到柱头上能够萌发,并同时引起柱头乳突状细胞的胼胝质形成(禾本科植物在花粉管顶部形成胼胝质)这是不亲和性的反应。种内不亲和与种间不亲和都发生这种反应,这种反应标志了两亲本之间尚有一定的亲缘关系;如果花粉既不萌发,又不引起乳突细胞呈现胼胝质反应,则反映了两亲本之间无亲缘关系的不可杂交性,这常是不同科间的远缘杂交表现的情况。
种间杂交的不亲和系统至今知道得很少,有一些实验证明花粉与柱头之间存在识别作用。银白杨和加拿大白杨的种间杂交通常是不育的,异己的花粉停滞在柱头上。但从银白杨的花粉提取出其表面的蛋白质与不亲和的加拿大白杨的花粉一起加在银白杨柱头上,则可获得一定数量的杂交种子;用杀死的银白杨花粉与加拿大白杨花粉混合后授与银白杨,也得到同样的结果。这表明母本花粉的壁蛋白与柱头的识别作用,可使不亲和的异己花粉被接受。这样的亲和花粉与异己花粉混合授粉的方法在生产上已作为产生杂种的一种手段。
受精_受精 -离体传粉和受精
20世纪60年代已可使被子植物带胎座的胚珠、整个雌蕊或子房置于离体培养的条件下实现传粉和受精。
1962年K.甘达及其合作者报道了他们用罂粟为材料在无菌条件下,将没有受精的带胎座的罂粟胚珠,置于装有消毒的营养培养基的试管中,同时用无菌针从花药中取出花粉撒在胚珠的表面上,结果发生了与正常相似的受精作用,最后结出有生活力的种子。这些种子可在试管中长成健康的幼苗,这是第一个离体传粉和受精成功的实验。迄今已报道有40多种植物获得成功,主要属罂粟科(如蓟罂、虞美人)、茄科(如烟草、矮牵牛)、石竹科(如麝香石竹、异株女娄菜)及十字花科(如甘蓝)的植物,近年来又在禾本科的玉米和小麦中获得成功。
离体传粉和受精的技术在育种过程中有可能作为克服有性生殖中自交或杂交存在的不亲和性障碍的手段。自交不亲和植物在受精过程中的障碍,通常发生在柱头或花柱中,即由于花粉与柱头识别反应导致花粉管生长受到抑制,不能进入柱头或在花柱中生长停滞以致受精失败。因此有可能利用离体的胚珠授粉顺利完成受精。这方面已在矮牵牛属植物试验成功。例如自交不亲和的腋花矮牵牛用一个子房的胎座上的全部胚珠进行离体培养和授以自花花粉。结果花粉管生长正常,最后形成具正常发育的胚和胚乳的种子。由离体受精得到的种子长成的第一代植株象它的亲本一样,是二倍体的,能育和自交不亲和的。另一方面,曾在石竹科、罂粟科、茄科及禾本科的一些植物试用离体传粉和受精的技术克服杂交不亲和性。其中许多杂交组合虽然能达到受精,但未能获得有生活力的种子。
不过也有少数得到杂种种子和幼苗的。如:①栽培烟草(2n=28)与德氏烟草或与莲座烟草(2n=40)杂交,这是常规的杂交方法难以成功的,然而,用离体传粉的方法,在试管里得到了杂种种子而且种子在培养基上萌发成幼苗;②玉米与墨西哥玉米在试管中杂交,获得了具发育良好的胚和胚乳的种子;③小麦(2n=42)的子房在离体培养条件下授以黑麦(2n=14)的花粉,得到具有器官分化的胚和含胚乳的颖果。将这种杂种胚从颖果中解剖出来进行胚培养,结果长成杂种幼苗。幼苗根尖染色体数为小麦和黑麦体细胞染色体半数之和,即28(21+7)个。