B淋巴细胞瘤-2基因简称bcl-2(B-cell lymphoma-2),是细胞凋亡研究中最受重视的癌基因之一。
bcl_bcl-2 -基因概述
自从1972年Kerr提出细胞凋亡(apoptosis)的概念至今,人们对细胞凋亡现象进行了广泛、深入的研究。但是,凋亡的分子和生化机制迄今尚未彻底明了,而已形成的初步认识大多源于对Bcl-2基因家族的研究。已知,凋亡进程可分为三个时相:诱导期,效应期和降解期。在诱导期,细胞接受各种信号从而引发各种不同的效应;进入效应期后,经过一些决定细胞命运(存活/死亡)的分子调控点,细胞进入不可逆的程序化死亡,这些调控分子包括一系列原癌基因和抑制癌基因的产生,其中Bcl-2家族起着决定性的作用;降解期则产生可见的凋亡现象。
Bcl-2基因(即B细胞淋巴瘤/白血病-2基因)是一种原癌基因,它具有抑制凋亡的作用,并且近年来的一些研究已开始揭示这一作用的机制。目前已经发现的Bcl-2蛋白家族按功能可分为两类,一类是像Bcl-2一样具有抑制凋亡作用,如哺乳动物的Bcl-XL、Bcl-W、Mcl-1、A1、线虫Ced-9、牛痘病毒E1B119kD等,而另一类具有促进凋亡作用,如Bax、Bcl-Xs、Bax、Bak、Bik/Nbk、Bid和Harakiri。
最初在血液淋巴细胞中发现Bcl-2能抑制细胞死亡,随后陆续在其它一些细胞中也发现Bcl-2的这种作用。但近年来研究发现,除这些之外尚存在Bcl-2不敏感的凋亡途径。
bcl_bcl-2 -敏感凋亡
Bcl-2可以抑制由多种细胞毒因素所引起的细胞死亡。Bcl-2的过度表达能增强所观察细胞对大多数细胞毒素的抵抗性。这一发现使人们认识到凋亡的各种信号转导途径有一个共同的通路或交汇点,而该通路或交汇点受Bcl-2调节。实验表明,Bcl-2可增强细胞对大多数DNA损伤因子的抵抗性,抑制大多数化疗药物所引起的靶细胞凋亡,但其本身并不能抑制这些因素对细胞的损伤;同样地,它也不能促进DNA修复。p53蛋白是DNA损伤的一个分子传感器,已证实Bcl-2能抑制p53介导的凋亡,但不能抑制p53向核内转位或者p53介导的生长停滞,可能Bcl-2的作用是在DNA损伤后,阻止激活凋亡机制的信号到达其靶分子;在细胞毒性T细胞中,由颗粒酶B激活Caspases家族的一个或多个半胱氨酸蛋白酶所诱导的凋亡不依赖于Bcl-2,颗粒酶B可能仅作用于凋亡路径中Bcl-2调节位点的下游。以上结果表明在凋亡途径中Bcl-2的作用位点是在信号分子和效应蛋白酶之间的位置。
Bcl-2抑制细胞凋亡可能与其以下几种作用有关:
① 细胞催抗氧化作用以往研究表明,在去除生长因子所诱发的凋亡中,活性氧损伤(ROS)是促发细胞死亡的主要诱因:Bcl-2的过度表达可减少氧自由基的产生和脂质过氧化物的形成。提示Cai等的实验表明Bcl-2的抗氧化作用是间接的,即可能在于抑制超氧阴离子的产生而不是直接清除活性氧。细胞色素c(cytochrome c, Cyt c)作为呼吸链中重要的电子传递体,它从线粒体内膜上的释放会阻断电子向下游传递体危及呼吸链的功能并导致超氧阴离子的加速产生;而Bcl-2可以抑制Cyt c的释放,从而抑制了超氧阴离子的产生。此外,Bcl-2还可以增加胞内的谷胱甘肽(GSH)等抗氧化剂,升高NAD/NADH的比值,抑制和凋亡相联系的GSH降低,促进GSH进入核内,从而影响细胞的氧化还原状态。但也有证据表明,在没有自由基的情况下Bcl-2也能抑制凋亡。因此,Bcl-2的性质远非仅仅是一种抗氧化剂。
② 抑制钙离子跨膜流动 Lam等曾报道钙泵特异性抑制剂Thapsigargen(TG)诱导的WEH17.2等细胞的凋亡可被Bcl-2所抑制,其原因是Bcl-2抑制了Ca2+的跨膜流动,提 示Bcl-2可通过调节胞内钙离子浓度来调节凋亡。然而Wei等在神经元细胞GT1-7中发现Bcl-2中发现Bcl-2可抑制由TG所诱导的凋亡,但并不能抑制TG诱导的胞内钙离子浓度升高;同时Jason等在中国仓鼠卵巢细胞5AHSmyc中去除细胞内贮存Ca2+后发现过度表达的Bcl-2仍可抑制凋亡,提示Bcl-2对胞内Ca2+浓度的调节作用也只能是其抑制凋亡的机制之一。
③离子通道蛋白和吸附/锚定蛋白最近Reed提出Bcl-2蛋白可能具有双重功能:离子通道蛋白和吸附/锚定蛋白,提出Bcl-2抑制凋亡的部分新机制。
③-1 离子通道蛋白 体外实验证实,Bcl-2、Bcl-X1和Bax能在线粒体上形成离子通道,提示它们可能参与调节一些与凋亡有关的细胞现象,如线粒体通透性改变(巨孔形成)和线粒体释放凋亡蛋白本科激活因子――Cyt c和凋亡诱导因子(AIF)。过度表达的Bcl-2能抑制线粒体通透性改变,并影响巨孔的形成,从而抑制凋亡。但也有实验发现即使线粒体通透性未改变,Cyt c等凋亡激蛋白也可以释放到胞液中从而诱发细胞凋亡,因此这种观点还有待进一步证实。
在所观察的细胞系及多种情况下,凋亡的发生都伴有Cyt c从线粒体释放,继而伴有Caspases激活,而活化的Caspases降解底物后,其蛋白降解产物又引起线粒体通透性改变,并最终导致凋亡发生。Yang等与Kluck等发现Bcl-2可以通过抑制线粒体释放Cyt c来抑制凋亡,继而Rosse等和Zhivotovsky等用两种不同的方法证明了即使Cyt c已经释放入胞浆,Bcl-2仍能延迟细胞死亡,这是由于Bcl-2抑制了活化的Caspases对线粒体释放的上游和下游都有Bcl-2调节凋亡的作用点。
AIF作为一种线粒体来源的效应蛋白酶,在分离在胞核中能诱导凋亡,同时还能激发线粒体通透性改变,诱导线粒体释放Cyt c和Caspase-9,并在体外切割和激活其底物――半胱氨酸蛋白酶CPP32。体外研究发现,Bcl-2过度表达可能阻止AIF从分离的线粒体上释放,但它并能影响AIF的产生,同时也不能影响AIF促凋亡的功能。
③-2 吸附/锚定蛋白 在正常情况下Bcl-2通过其C端疏水残基的伸展而锚定在细胞内膜上,作为吸附/锚定蛋白,可把胞液蛋白固定在膜边,或是引导胞液蛋白与别的膜蛋白相互作用。
Bcl-2可与Bcl-2家族的Bcl-X1、Bcl-Xs、Bax、Bcl-2、Bad和Mc1-1形成同源的蛋白二聚体,而特定的蛋白二聚体则可作为在细胞死亡信号通路上的分子开关。例如,Bcl-2可与促凋亡Bax形成二聚体,如果Bax相对量高于Bcl-2,则Bax同二聚体的数量增多,从而促进细胞死亡;而如果Bcl-2相对量高于Bax,则促进形成Bcl-2/Bax异二聚体,并使Bcl-2同二聚体的量增多,从而抑制细胞死亡;而促凋亡分子和Bad和Bcl-Ss则竞争结合细胞死亡抑制分子如Bcl-X1或Bcl-2,由此替换了Bax并促进Bax同二聚体形成,诱导凋亡。
另外,Bcl-2还可与Bcl-2家族外的11种蛋白质结合,包括蛋白激酶Raf-1、蛋白磷酸酶Calcineurin、GTP酶、R-Ras和H-Ras、p53-BP2、朊蛋白Pr-1、Ced-4、BAG-1、Nip-1、Nip-2和Nip-3,并由此实现其抗凋亡作用。
bcl_bcl-2 -不敏感
Bcl-2抑制凋亡的作用并不是万能的,例如在细胞毒T细胞杀伤作用中,特定的淋巴因子撤离以及某些TNF受体家族介导的凋亡途径中Bcl-2不能抑制凋亡,其原因右能是这些凋亡诱导剂作用于Bcl-2下游或是独立于Bcl-2的其它凋亡路径。最近Scaffidi等发现,Jurkat等细胞(Ⅱ型细胞)中Bcl-2可抑制TNF受体家族中的CD95(Apo-1/Fas)信号通路介导的凋亡,而在SKW6.4等细胞(Ⅰ型细胞)中Bcl-2则不能抑制这种信号通路介导的凋亡。其结果表明,凋亡途径对Bcl-2敏感与否可能与细胞类型有关。进一步研究发现,这种差异取决于线粒体是否参与这种凋亡途径,在Ⅰ型 细胞中线粒体则未参与该凋亡途径。因此,Bcl-2可能只抑制依赖于线粒体的凋亡途径。
更进一步研究发现,来源于牛痘病毒的细胞因子应答修饰体A(crmA)是一种丝氨酸蛋白酶抑制剂(serpin)类分子,能抑制一些Caspases的功能。而来源于杆状病毒的P35蛋白也能抑制一些Caspases的功能,而且它比CrmA具有更广泛的Caspases抑制特性。在组织培养体系中或转基因鼠的T淋巴细胞中CrmA表达能抑制CD95(Fas/Apo-1)和p55TNF受体Ⅰ诱发的凋亡,但对另一些诸如生长因子撤除,DNA损伤等细胞毒因素引起的凋亡则无能为力,而这些由细胞毒因素引起的凋亡现象则可被Bcl-2及其同系物所抑制,但是Bcl-2又不能抑制由CD95(Fas/Apo-1)和TNF受体介导的凋亡。由细胞毒因素及TNF受体家族介导的凋亡都可有效地被p35所抑制,提示这些通路都依赖于Caspases的激活。上述结果表明,在细胞中不同的凋亡激活途径同时存在,其一是需要对CrmA敏感的Caspases参与但又不能被Bcl-2所抑制;而另一个则是Bcl-2可抑制的凋亡途径,其中Caspases对p35敏感,但对CrmA则不敏感。
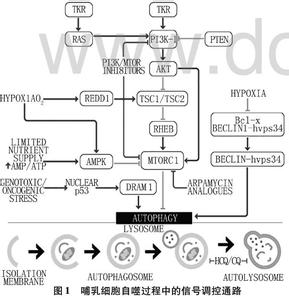
bcl_bcl-2 -总结
目前Bcl-2是凋亡分子机制研究的主要靶分子。随着对Bcl-2以及凋亡本身研究的日渐深入,Bcl-2的作用机制和凋亡的分子机制最终被阐明,从而提高对与凋亡有关的疾病的认识和诊治水平。
B-cell CLL/lymphoma 2
在哺乳动物细胞中,调节线粒体外膜通透性,大多数定位在线粒体外膜上,或受到信号刺激后转移到线粒体外膜上。根据其功能可分为两组:Bcl-2, Bcl-xl, Bcl-w等抑制细胞凋亡;Bax,Bak,Noxa等促进细胞凋亡。
关于Bcl-2家族调控线粒体外膜通透性的机制,假说之一是,细胞接受凋亡信号后促凋亡因子Bax和Bak发生寡聚化,从细胞质中转移到线粒体外膜上,并与膜上的电压依赖阴离子通道相互作用,使通道开放到足以使线粒体内的凋亡因子如细胞色素c等释放到细胞质基质中,引起细胞死亡。




