生殖细胞是多细胞生物体内能繁殖后代的细胞的总称,包括从原始生殖细胞直到最终已分化的生殖细胞。此术语由A・恩格勒和K・普兰特尔于1897年提出以与体细胞相区别。体细胞最终都会死亡,只有生殖细胞有延存至下代的机会。物种主要依靠生殖细胞而延续和繁衍。长期的自然选择使每一种生物的结构都为其生殖细胞的存活提供最好的条件。
生殖_生殖细胞 -概述
生殖细胞生殖细胞(germcell)是多细胞生物体内能繁殖后代的细胞的总称,包括从原始生殖细胞直到最终已分化的生殖细胞。此术语由A・恩格勒和K・普兰特尔于1897年提出以与体细胞相区别。体细胞最终都会死亡,只有生殖细胞有延存至下代的机会。物种主要依靠生殖细胞而延续和繁衍。长期的自然选择使每一种生物的结构都为其生殖细胞的存活提供最好的条件。在个体发生中,生殖细胞在发育早期就被决定了。在一些动物中,其决定因子可以追溯到上一代卵细胞的卵质。例如果蝇卵的后部有一特殊的细胞质区域称极质,其中富含核糖核酸(RNA)的小颗粒叫极颗粒。经过受精、卵裂,含有极颗粒的细胞称极细胞,就是果蝇的原始生殖细胞。如果将原在后部的极质注射到卵的前部,可使预定发育为体细胞的细胞发育为生殖细胞。
生殖_生殖细胞 -简介
在单细胞生物群体中已有生殖细胞分化的迹象。如团藻在大多数小形营养细胞间出现了少数大的生殖细胞。在个体发生中,生殖细胞在发育早期就被决定了。在一些动物中,其决定因子可以追溯到上一代卵细胞的卵质。例如果蝇卵的后部有一特殊的细胞质区域称极质,其中富含RNA的小颗粒叫极颗粒。经过受精、卵裂,含有极颗粒的细胞称极细胞,就是果蝇的原始生殖细胞。如果将原在后部的极质注射到卵的前部,可使预定发育为体细胞的细胞发育为生殖细胞。在哺乳动物胚胎中生殖细胞被决定后,需通过迁移到达生殖腺的部位并在那里分化。生殖_生殖细胞 -产生
昆虫的生殖细胞
施旺和许耐登来说细胞仍然主要是一种结构成分,但在19世纪功年代其他学者已经强调了细胞的生理功能,尤其是发育和营养功能。随着对细胞及其组成成分(特别是细胞核)的了解日益深入,“细胞学说”这一概念的意义也逐渐发生了变化。许耐登的学说直接促进了区分动植物细胞的非常活跃的研究工作。1852年Remak(1815-1865)指陈,蛙卵是细胞,在发育中的蛙胚中新细胞是由原已存在的细胞分裂产生的。他着重驳斥了自由细胞形成学说。在这一点上他得到了微尔科(1855)的支持,后者指出,就许多正常的和病态的动物及人类组织来看,每个细胞都是从原已存在的细胞分裂而来。他断言:“作为一般原则,任何种类的发育都不是从头开始,因而(必须)否定(自然)发生学说,不论是在个别部分的发育史上还是在整体生物的发育史上”(Virchow,1858:54)。Kolliker以及其他一些植物学家也在同一时期得出了同样的结论,然而,由于许耐登的权威性而延迟了这结论在植物学界被普遍接受。达尔文在1868年(II:370)还拿不准自由细胞形成究竟是否可能。后来微尔科的格言“细胞来自(原先存在的)细胞”,(Omniscellula e cellula)虽然终于被每个人接受,然而那个时候对细胞(尤其是核)分裂过程的细节还并不清楚(见下文,“有丝分裂”)。
由于有了对细胞的这种新的解释,重新考虑受精过程的时机已经成熟。如果躯体的一切部位都由细胞组成,则性腺(卵巢和睾丸)是否也是如此?雄性和雌性的“种子物质”究竟是什么?它是否也由细胞构成?雌、雄性的性细胞有什么区别?这一类早已构思过的问题当然并不是一开始就提了出来,然而这些问题必然是细胞学说的逻辑结果。后来越来越明显只有等到阐明了受精过程中细胞的作用,真正有生命力的遗传学说才能建立。正是在这几十年中生殖细胞这个概念才脱颖而出。
生殖_生殖细胞 -形成条件
多细胞生物体内能繁殖后代的细胞的总称,包括从原始生殖细胞直到最终已分化的生殖细胞。此术语由A.恩格勒和K.普兰特尔于1897年提出以与体细胞相区别。体细胞最终都会死亡,只有生殖细胞有延存至下代的机会。物种主要依靠
生殖细胞而延续和繁衍。长期的自然选择使每一种生物的结构都为其生殖细胞的存活提供最好的条件。
在单细胞生物群体中已有生殖细胞分化的迹象。如团藻科的杂球藻有4个较小的细胞失去分裂能力,专司运动和代谢,称为营养个体,其余28个有分裂能力,称为生殖个体;团藻则在大多数小形营养细胞间出现了少数大的生殖细胞。
在个体发生中,生殖细胞在发育早期就被决定了。在一些动物中,其决定因子可以追溯到上一代卵细胞的卵质。例如果蝇卵的后部有一特殊的细胞质区域称极质,其中富含RNA的小颗粒叫极颗粒。经过受精、卵裂,含有极颗粒的细胞称极细胞,就是果蝇的原始生殖细胞。如果将原在后部的极质注射到卵的前部,可使预定发育为体细胞的细胞发育为生殖细胞。在马副蛔虫卵中也可以看到类似的情况。有证据表明,两栖类生殖细胞可能也是以同样的方式决定的。但还不了解在哺乳动物胚胎中,是什么因素决定某些细胞发育成生殖细胞的。只知道生殖细胞被决定后,需通过迁移到达生殖腺的部位并在那里分化(见生殖质)。
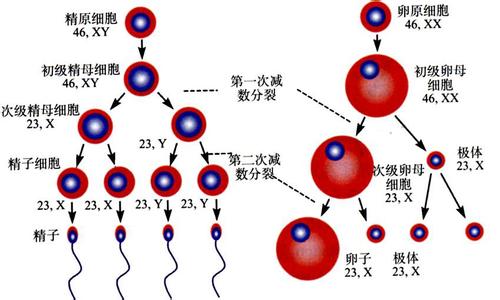
生殖_生殖细胞 -结合
精子的获能 精子中的半数含Y染色体(23,Y),半数含X染色体(23,X)。射出的精子虽有运动能力,却无穿过卵子周围放射冠和透明带的能力。这是由于精子头的外表有一层能阻止顶体酶释放的糖蛋白。精子在子宫和输卵管中运行过程中,该糖蛋白被女性生殖管道分泌物中的酶降解,从而获得受精能力,此现象称获能(capacitation)。精子在女性生殖管道内的受精能力一般可维持1天。
卵子的成熟 从卵巢排出的卵子处于第二次成熟分裂的中期,并随输卵管伞的液流进入输卵管,在受精时才完成第二次成熟分裂。若未受精,于排卵后12~24小时退化。
生殖_生殖细胞 -分类
生殖细胞可以分成孢子和配子两类。孢子是不需配合的生殖细胞,通常是无性的,可由减数分裂或有丝分裂产生,见之于原生动物中的孢子虫纲和植物中。
配子是需经配合成合子后方能发育的生殖细胞,也称性细胞,由减数分裂或有丝分裂产生。产生配子的细胞称配子母细胞。这是未分化的原始生殖细胞,可在雄性或雌性生殖腺中分别分化为精子和卵。在低等植物中,配子母细胞在大多数情况下直接构成称作配子囊的性器官。例如在原植体植物(Thallophyte)中的同形配子囊产生同形配子;异形配子囊产生异形配子等。
生殖_生殖细胞 -受精
动、植物受精作用的实质是父本和母本的生殖细胞(配子)互相融合,这两个配子在形成新的合子上各自作出了同样贡献,而且关键过程是两个配子的细胞核相互融合,这些观点到了1844年左右已经逐步确立并被有关学者普遍接受。人们的注意力便开始转移到细胞核上。细胞核是不是就像后生论者所设想的,仅仅是一团无定形的胚样物质、也许只是在融合时才激发了卵细胞的发育过程?或者是细胞核虽小,却具有严密结构,这肉眼不可见的显微结构是否就是受精作用之后的一切非常精确并具有特异性的发育过程的关键?如果把细胞核仅仅看作是细胞发育和细胞分裂的引发物,就会认为它在完成了这一任务后就会被溶解掉,在新的细胞分裂之前或至少是在配子形成之前再重新形成。由于19世纪后半期的细胞学家所接受的都是生理学家或胚胎学家的教育训练,他们的侧重点是发育问题,因而用不着关心细胞核的连续性。他们很少过向性状是怎样从亲代传递到子代的遗传学问题。
1875-1880年之间由于Balbiani,van Beneden,Flemming。Schleicher,Strasburger等五位学者在细胞分裂过程中能够不断追踪其全部进程,因而“自由细胞形成”或细胞核“重新”形成这种信念的最后残余才得以完全肃清。这几位学者论证了。
三项重要事实:
(1)在细胞分裂之前细胞核开始分裂,
(2)细胞核物质的变化具有正常顺序(见下文),
(3)细胞核分裂和细胞分裂的基本现象在植物界和动物界都是相同的。
越来越明显的是,细胞核的作用不单是生理性的(即纯粹物理意义上的作为细胞分裂的引发物)。它是具有严密组织的、很可能是按特定格局构成的结构。这种格局的实质此后一直是细胞学者不断关心的问题,仍然没有求得最后答案。关于这方面研究进展的特点是分析越来越细。所涉及的步骤是从完整个体转向细胞,从整个细胞转向细胞核,现在则从完整的核转向到它的主要结构成分、染色体。
生殖_生殖细胞 -鱼类
有圆形外观、偏心的核、以及充满颗粒的细胞质,这些都是原始生殖细胞所具有的典型之形态特征。鱼类的第一个代理孕母。然而,为了获得具有功能的配子,仍需努力把原始生殖细胞转变成为精子和卵子的技术找出来。日本东京海洋大学的吉崎悟朗教授及其研究团队曾以樱鲑为代理孕母,在樱鲑的腹膜腔内移植虹鳟的原始生殖细 还是多产的物种,只需一次受精,就可获得数以千计,含有绿色萤光蛋白标记的原始生殖细胞的转殖基因胚。一般而言,在鱼类生殖隆起中的原始生殖细胞的数目很少,若要进行移植或体外培养实验时,多产就成为一个很有利的条件,而虹鳟正好具有这个优点。流式细胞仪可以用来纯化被绿色萤光蛋白标记的原始生殖细胞。从孵化期的胚上切下来的生殖隆起,经胰蛋白酉每溶解分散,透过流式细胞仪可分类成为被绿色萤光蛋白标记的细胞,和不被绿色萤光蛋白标记的细胞。前者直径大约20微米,是一种具胞,藉由这个方法从樱鲑生产虹鳟子代,这是世界上鱼类的第一个代理孕母研究成功的报导。又为追踪外源的原始生殖细胞在代理孕母体内的表现,研究人员先利用绿色萤光蛋白基因,把原始生殖细胞标记,然后再把孵化胚生殖隆起内的原始生殖细胞植入代理孕母。被植入到代理孕母腹膜腔内的原始生殖细胞,会朝向代理孕母的生殖隆起移动并形成群体聚落。到后来,来自供核者虹鳟的原始生殖细胞,在不同种(樱鲑)的代理孕母的生殖腺中增殖,并分化为成熟的卵子和精子,再透过这些制造的配子受精,可生产出许多来自供核者的活仔鱼。