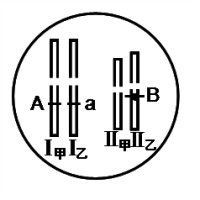
染色体组分析是对异源多倍体植物的染色体组来源进行的分析。方法主要是将异源多倍体植物与假定的基本种杂交,然后观察杂交子代在减数分裂过程中染色体的配对行为。
染色体组_染色体组分析 -染色体组分析
染色体组_染色体组分析 -正文
对异源多倍体植物的染色体组来源进行的分析。方法主要是将异源多倍体植物与假定的基本种杂交,然后观察杂交子代在减数分裂过程中染色体的配对行为。在减数分裂中,同源染色体通过配对(联会)形成二倍体,非同源染色体因不能联会而呈单倍体状态。如果异源多倍体植物和基本种的杂交子代的减数分裂过程中出现相当于基本种染色体基数的二倍体,便说明异源多倍体的一个染色体组来源于这一基本种。某些基因能干扰染色体的配对,从而给二倍体分析带来困难或错误。英国细胞遗传学家R.赖利等在60年代中发现小麦 5B染色体的长臂上有一个基因 ph,它使部分同源染色体的联会受到阻碍。在拟山羊草(Aegilopsspeltoides)中还有阻碍作用更大的基因。在玉米和小麦中发现的不联会基因可以使同源染色体在减数分裂中以单价体形式出现。因此,在染色体组分析中还常采用一些辅助的方法,包括解剖学、组织学、形态学、生物化学(特别是同工酶分析)的方法。
染色体组分析有助于对物种起源的了解,也可以为倍性育种提供参考资料。
美国细胞遗传学家T.H.古得斯皮德和R.E.克劳森在1928年首先用二倍体分析方法研究了具有48个染色体的栽培烟草(Nicotiana tabacum)的起源。最初根据形态特征认为它的祖先是两种二倍体野生烟草:林烟草(N.sylvestris,2n=24)和绒毛烟草(N.tomentosa,2n=24)。染色体组分析结果说明栽培烟草与二者分别杂交得到的子代在减数分裂中都只出现12个二倍体,说明栽培烟草与这两个二倍体物种间都有一个染色体组是相同的。但是林烟草与绒毛烟草的杂交子代在减数分裂中却出现24个单价体,说明它们的染色体组是完全不同的。这一杂种染色体加倍后得到的异源四倍体植株虽然在形态上很像栽培烟草,但不结种子,因此可以判定单由这两个物种不能形成栽培烟草。而后才发现与绒毛烟草相近的另外两个二倍体物种绒毛状烟草 (N.tomentosiformis)和耳状烟草(N.otophora)分别与林烟草形成的异源四倍体都是可育的,它们才是栽培烟草的祖先。从进化的角度来说,四倍体烟草属于复合种,构成复合种的祖先二倍体种称为基本种。
1930年日本遗传学家木原均首先提出染色体组分析这一名词,并在1930~1951年间和他的同事一起对小麦属(Triticum)和山羊草属(Aegilops)进行了详细的染色体组分析。他们根据染色体组的异同,将小麦属的20多个种分为四大类:①具有7对A组染色体的二倍体一粒小麦系(T.Einkorngroup);②具有14对A、B组染色体的四倍体二粒小麦系(T.Emmergroup);③具有 21对A、B、D组染色体的六倍体普通小麦系(T.Dinkelgroup);④具有14对A、G组染色体的四倍体提摩菲维小麦系(T.Timopheevigroup)。组成小麦属各个种的四个染色体组中只有 A和 D的来源是肯定的。A组来自一粒小麦(T.monococcum),D组来自山羊草属的节节麦(A.squarrosa),B组可能来自拟山羊草,G组究竟来自什么物种还缺乏证据。后来用其他分析方法说明节节麦和尾形山羊草(A.caudata)可能是异源多倍体具节山羊草(A.cilindrica)的祖先。同工酶的分析结果指出后者的电泳图谱和前两者的混合物的电泳图谱相同,验证了上述结论。
染色体组分析还可以为倍性育种提供参考资料。例如白菜型油菜(Brassicacampestris)有10对染色体,属A组;甘蓝(B.oleracea)有9对染色体,属C组;黑芥(B.nigra)有8对染色体,属B组。A组与C组所形成的异源四倍体就是具有19对染色体的欧洲油菜(B.napus);A组与B组形成具有18对染色体的芥菜型油菜(B.juncea);B组与C组形成具有17对染色体的埃塞俄比亚油菜(B.carinata)。
日本学者禹长春根据染色体组分析结果,用白菜型油菜与甘蓝杂交,并使杂种染色体数目加倍,合成了一个与欧洲自然油菜相似的异源四倍体,这一育种措施有利于把某些白菜和甘蓝类中的优良特性引入到油菜中去(见染色体倍性)。
参考书目
木原均:《小