感悟:随着科技的进步,对事物的认识会更完整和科学。1930—1950年,随着电子显微镜技术的发展,当应用这项技术来研究细胞时,才发现细胞的边界膜是一个固体结构的实体,从而证实了细胞膜的存在。
17世纪中叶以后的2个世纪中,细胞学说的发展史已经大体完成。但是唯独对细胞膜的认识还要推迟两个世纪。
1855年,耐格里发现色素透入已损伤和未损伤的植物细胞的情况并不相同。他便通过细胞的渗透特性去研究它的“边界”。耐格里和克拉默一起进行实验,通过实验发现细胞具有敏感的渗透特性,它的体积可以随着周围介质的不同渗透强度而改变。当细胞外面的溶质渗透强度大时,细胞就变小;溶质渗透强度小时,细胞就变大。耐格里提出,细胞与环境之间正是通过这种“边界”发生关系的。
德国植物生理学家普费弗对植物细胞的渗透行为进行了大量的试验,并于1897年提出了两个重要的结论:第一,细胞是被质膜包被着的;第二,这层质膜是水和溶质通过的普遍障碍。同时,很快又发现,细胞膜这个屏障具有明显的选择性,一些物质可通过它,而另一些物质几乎完全不能通过。
E. Overton 1895 发现凡是溶于脂肪的物质很容易透过植物的细胞膜,而不溶于脂肪的物质不易透过细胞膜,因此推测细胞膜由连续的脂类物质组成。

水溶性物质难以通过质膜
1925年E.Gorter & F. Grendel 用有机溶剂提取了人类红细胞质膜的脂类成分,将其铺展在水面,测出膜脂展开的面积二倍于细胞表面积,因而推测细胞膜由双层脂分子组成。
以下就是研究细胞膜的过程中,科学家建立的结构模型。
1.片层结构模型
1935年J. Danielli & H. Davson发现质膜的表面张力比油-水界面的张力低得多,推测膜中含有蛋白质,从而提出了”蛋白质—脂类—蛋白质”的三明治模型。认为质膜由双层脂类分子及其内外表面附着的蛋白质构成的。
1954年对该模型进行了修改:膜上有一些二维伸展的孔,孔的表面也是由蛋白质包被的,这样使孔具有极性,可提高水对膜的通透性。
这一模型是第一次用分子术语描述的结构,并将膜结构同所观察到的生物学理化性质联系起来,对后来的研究有很大的启发。
2.“单位膜”模型
1959 年J.D. Robertson用超薄切片技术获得了清晰的细胞膜照片,显示暗—明—暗三层结构,厚约7.5nm。这就是所谓的“单位膜”模型。它由厚约3.5nm的双层脂分子和内外表面各厚约2nm的蛋白质构成。
单位膜模型是在片层结构模型的基础上发展起来的另一个重要模型。它与片层结构模型有许多相同之处,最重要的修改是膜脂双分子层内外两侧蛋白质存在的方式不同。单位膜模型强调的是蛋白质为单层伸展的β折叠片状,而不是球形蛋白。另外,单位膜模型还认为膜的外侧表面的膜蛋白是糖蛋白,而且膜蛋白在两侧的分布是不对称的。
这一模型能够解释细胞质膜的一些基本特性,例如质膜有很高的电阻,这是由于膜脂的非极性端的碳氢化合物是不良导体的缘故;再如由于膜脂的存在,使它对脂溶性强的非极性分子有较高的通透性,而脂溶性弱的小分子则不易透过膜。
单位膜模型的不足之处在于把膜的动态结构描写成静止的不变的。该模型也不能解释为什么有的膜蛋白很容易被分离,有些则很难。
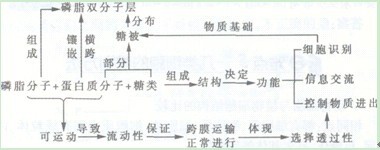
3.流动镶嵌模型
1972年Singer和Nicolson根据免疫荧光技术、冰冻蚀刻技术的研究结果,在“单位膜”模型的基础上提出“流动镶嵌模型”。认为球形膜蛋白分子以各种镶嵌形式与脂双分子层相结合,有的附在内外表面,有的全部或部分嵌入膜中,有的贯穿膜的全层,这些大多是功能蛋白。
流动相嵌模型有两个主要特点。其一,蛋白质不是伸展的片层,而是以折叠的球形镶嵌在脂双层中,蛋白质与膜脂的结合程度取决于膜蛋白中氨基酸的性质;其二,膜具有一定的流动性,不再是封闭的片状结构,以适应细胞各种功能的需要。
这一模型强调了膜的流动性和不对称性,较好地体现细胞的功能特点,被广泛接受,也得到许多实验的支持。后来又发现碳水化合物是以糖脂或糖蛋白的形式存在于膜的外侧表面。
4.晶格镶嵌模型
流动镶嵌模型的补充,强调流动的整体性。1975年,Wallach提出了晶格镶嵌模型。他在流动镶嵌模型的基础上,进一步强调:生物膜中流动性脂质的可逆性变化。这种变化区域呈点状分布在膜上。相变表现为膜脂分子的一种协同效益,即几十个以上的脂分子同时相变。膜脂的相变受温度、脂本身的性质、膜中其他成分、pH和二价阳离子浓度等因素的影响。
5.板块镶嵌模型
1977年,Jain和White提出了板块镶嵌模型,其内容本质上与晶格镶嵌模型相同。他们认为:在流动的脂双分子层中,存在许多大小不同的、刚度较大的、彼此独立运动的脂质“板块”(有序结构区),板块之间被无序的流动的脂质区所分割,这两种区域处于一种连续的动态平衡之中。
6.脂筏模型
脂筏模型(lipidrafts model)即在生物膜上胆固醇富集而形成有序脂相,如同脂筏一样载着各种蛋白。脂筏是质膜上富含胆固醇和鞘磷脂的微结构域。大小约70nm左右,是一种动态结构,位于质膜的外小页。由于鞘磷脂具有较长的饱和脂肪酸链,分子间的作用力较强,所以这些区域结构致密,介于无序液体与液晶之间,称为有序液体。在低温下这些区域能抵抗非离子去垢剂的抽提,所以又称为抗去垢剂膜。脂筏就像一个蛋白质停泊的平台,与膜的信号转导、蛋白质分选均有密切的关系。
从脂筏的角度来看,膜蛋白可以分为三类:①存在于脂筏中的蛋白质;包括糖磷脂酰肌醇锚定蛋白,某些跨膜蛋白等;②存在于脂筏之外无序液相的蛋白质;③介于两者之间的蛋白质,如某些蛋白在没有接受到配体时,对脂筏的亲和力低,当结合配体,发生寡聚化时就会转移到脂筏中。
脂筏中的胆固醇就像胶水一样,它对具有饱和脂肪酸链的鞘磷脂亲和力很高,而对不饱和脂肪酸链的亲和力低。