类别:人类学
讨论古代世界历史和文明起源问题,需要人类学的背景知识。特推荐以下两篇论文供博友参考:
一、华夏人类的起源
【体质人类学基本知识】
在本文的一开始,首先需要纠正大家的一个传统观念:中国人皮肤的颜色不是黄色的,而是浅棕色的。中国人眼睛的颜色不是黑色的,而是深棕色的。中国人的头发的颜色也不是纯黑色的,而是带有一点深棕色的。对于中国人种族上的这种性状特征,我们需要了解它的来源。在此之前,我们又必须了解中国人的种族构成。
中国人的种族来源,由两大部分构成,一部分是欧亚部落的血统,一部分是古亚洲人的血统。古亚洲人是第一次出非洲的人群,而欧亚人是第二次出非洲的人群中的第二次扩张的人群。
古亚洲人分为两种,一种是矮黑人(D-YAP),他们和非洲黑人(尼格罗人)拥有一个共同的祖先,另一种是棕种人(C-M130)。纯种的矮黑人是印度的安达曼人,东南亚的维达人等,而纯种的棕种人如今已不存在,但他们是蒙古人,女真人,澳大利亚土著等民族的直系祖先。矮黑人,棕种人先后从亚洲南部北上东亚后,肤色变浅。矮黑人中最进化的一支是日本的阿伊努人(虾夷人),它们是绳文人的后裔,也是大和民族的重要底子。另外,西藏人中也拥有大量的矮黑人父系成分。
第二次出非洲的人群被称为中东部落(F-M89),其中的一支演化为欧亚部落(K-M9),其余进化成地中海-高加索人种,属于暗白人种。欧亚部落的原始人种属于未分离的黄白人种,又演化成好几支。其中一支形成东亚部落(NO-M214),被称为黄种人(又称华夏-芬兰人种),另有一支形成中亚部落,属于早期的白种人。
东亚部落的黄种人又分离为两支,一支是芬兰人,部分北亚人的直系祖先(N-M231)。另一支则形成中国人,东南亚人的直系祖先(O-M175)。在M175的基础上,东亚地区的黄种人演化为华夏,东夷,百越,苗瑶,百濮,南岛等一系列民族。
而中亚部落也分化为两支,一支是印第安人和他们的北亚祖先(Q-P36),一支是主流欧洲人和印度雅利安人的祖先(R-M207)。其中进入欧洲的白种人是后来的日耳曼人,斯拉夫人,波罗的人,克尔特人的主要祖先(当然这些民族还有少部分其他来源)。这些生活在欧洲北部的人群是相对比较纯的欧亚部落后裔(主要是中亚部落,少数属于东亚部落),他们被称为诺迪克人种(又称北欧人种,白海-波罗的人种),属于显白人种,也是狭义上的白种人。他们和南欧的地中海人属于不同的人种。
在混血程度不高的史前,相比于纯种的地中海人和阿拉伯人,纯种的诺迪克—雅利安人和东亚的纯种黄种人血缘关系要更接近。这一点非常重要,也是下文讨论的基础。
北欧的诺迪克人种和南欧的的地中海人种在欧洲中部地区发生混血,形成了阿尔卑斯人种。一般认为,诺迪克人是典型的白种人,而阿尔卑斯人是典型的欧洲人,地中海人则接近中东人。亚洲黄种人和亚洲棕种人矮黑人大约在10万年前就已经分离,而亚洲黄种人和北欧白种人大约在4万年前的伊朗高原上分离,而传统认为东亚的黄种人和欧亚大陆北部的黄种人大约在3.5万年在阿富汗分离,不过最新的研究显示这两者的实际分离时间可能比原来的认识大大缩短,并且分离地点很可能在云南一带。另外需要说明,中国人也有部分芬兰-乌拉尔黄种人的血统,不过和欧洲黄种人属于不同的子类型。这些也非常的重要。回到中国,我们简要得概括一下中国人大体上的血缘构成:
父系=压倒性的黄种男人(F系下的K系)+极少量矮黑男人(M168下的C)+极少量棕种男人(M168下的D)+极少量的暗白种男人(M168下的F系)+极少量的显白种男人(F系下的K系)((黄种成分南北差异不大))
母系=部分黄种女人(N系下的R系)+部分矮黑女人(非非洲人的M系)+部分棕种女人(非非洲人的M系)+少量暗白种女人(非非洲人的N系)+可能的极少量的显白种女人(N系下的R系)((黄种成分由南向北递减))
以上是以父系的Y染色体遗传标记(父传子不传女,子传孙)和母系的mtDNA遗传标记(母传子女,女传后代,但子不能传后代)为标准的大致比例构成。但决定体质的主要是常染色体。常染色体是一个人父亲家族的所有直系祖先和母亲家族的所有直系祖先的混合,但比较容易被环境改变而不太稳定。中国人身上具体到底有多少是欧亚人血统,有多少是古亚洲人血统,这是一个很复杂的,至今还未有定论的问题。但可以肯定南方人是比北方人更纯的黄种人,中国最纯的黄种人是在两广一带,这些是没有任何问题的。虽然南北汉族父系有80%以上的同宗,母系也有50%的同宗,但是从各种方法研究常染色体的结果来看,北方汉族和北方少数民族,日韩人棕种成分比较多,并且聚类在一起,南方汉族和东南亚人的黄种成分比较多,并聚类在一起,两者之间以长江为界径渭分明。这说明南方人总体上的的黄种血统普遍要多于棕黑血统,而北方人则不好说。(其实这个结论从母系遗传的线粒体DNA各类型的分布情况就可以得出了)。由于历史上北方汉族的大规模南下,使得今天的南方汉人融入了更多的古亚洲血统,和东南亚人区分了开来。当然,气候也能影响常染色体的反映状况,这种聚类也有气候的因素。
在开始分析中国人的相貌特点以前,有必要列举新亚洲人(纯黄种人),古亚洲人(矮黑棕种人),诺迪克人(北欧人),地中海人(包括中东人)之间的一些体质上的性状特征如下:
1.纯黄种人 棕色眼睛 双眼皮 大眼睛 两眼居中 中眼窝 中鼻梁 阔鼻头 瓜子脸 面部线条柔和 深棕发 直发 长头型 中颧骨 浅棕皮肤 身材中等修长 无罗圈腿 体毛较少 脑容量最大
2.古亚洲人 黑色眼睛 单眼皮 小眼睛 两眼靠鼻 浅眼窝 扁鼻梁 塌鼻头 驴窄脸 面部无立体感 纯黑发 卷发 短头型 高颧骨 深色皮肤 身材矮小粗壮 有罗圈腿 体毛发达 脑容量一般
3.诺迪克人 蓝灰眼睛 双眼皮 大眼睛 两眼居中 中眼窝 中鼻梁 圆鼻头 介于14 面部线条柔和 金黄发 直发 长头型 中颧骨 白色皮肤 身材高大强壮 无罗圈腿 体毛中等 脑容量较大
4.地中海人 棕褐眼睛 双眼皮 大眼睛 两眼居中 深眼窝 高鼻梁 鹰钩鼻 椭圆脸 面部立体感强 棕褐发 卷发 长头型 高颧骨 浅棕皮肤 身材中等较壮 无罗圈腿 体毛发达 脑容量一般
需要说明的是,人类学中一般不用眼睛,皮肤,头发的颜色作为判定种族的根本依据,因为这些在史前的进化过程中是很容易改变的。即使在进化的自然选择作用大大降低的现代社会,一个民族的高矮胖瘦也是比较容易改变的。所以这些都不是根本性的人种特征。判定种族差异的根本区别主要在于人的骨骼方面,尤其是测定各项头骨数据,因为骨骼尤其头骨比较稳定不容易被环境改变。这样,我们可以发现,除去某些处于边缘化的印第安人在北亚的近亲部落以外(北欧人与地中海人混血而成的阿尔卑斯人是后来形成的),东亚的纯黄种人和北欧印度的纯白种人其实是整个欧亚大陆上血统最接近的人种!这不仅在已经遗传科学上获得了证明,而且在体质观察中也可以被确认。
写到这里,积累了上面的这些基础知识,现在就可以着手解决标题所提出问题:既然北欧人的祖先和中国人的祖先血缘关系如此接近,为什么中国人长得和北欧人不一样呢?为什么中国人没有强壮的体格,高鼻深目,金发碧眼,白色的皮肤呢?
我国人的种族来源,由两大部分构成,一部分是欧亚部落的血统,一部分是古亚洲人的血统。古亚洲人是第一次出非洲的人群,而欧亚人是第二次出非洲的人群中的第二次扩张的人群。古亚洲人分为两种,一种是矮黑人(D-YAP),他们和非洲黑人(尼格罗人)拥有一个共同的祖先,另一种是棕种人(C-M130)。纯种的矮黑人是印度的安达曼人,东南亚的维达人等,而纯种的棕种人如今已不存在,但他们是蒙古人,女真人,澳大利亚土著等民族的直系祖先。矮黑人,棕种人先后从亚洲南部北上东亚后,肤色变浅。矮黑人中最进化的一支是日本的阿伊努人(虾夷人),它们是纹绳人的后裔,也是大和民族的重要底子。另外,西藏人中也拥有大量的矮黑人父系成分。第二次出非洲的人群被称为中东部落(F-M89),其中的一支演化为欧亚部落(K-M9),其余进化成地中海-高加索人种,属于暗白人种。欧亚部落的原始人种属于未分离的黄白人种,又演化成好几支。其中一支形成东亚部落(NO-M214),被称为黄种人(又称华夏-芬兰人种),另有一支形成中亚部落(P*),属于早期的白种人。东亚部落的黄种人又分离为两支,一支是芬兰人,部分北亚人的直系祖先(N-M231),另一支则形成我国人,东南亚人的直系祖先(O-M175)。在M175的基础上,东亚地区的黄种人演化为华夏,东夷,百越,苗瑶,百濮,南岛等一系列民族。而中亚部落也分化为两支,一支是印第安人和他们的北亚祖先(Q-P36),一支是主流欧洲人和印度雅利安人的祖先(R-M207)。其中进入欧洲的白种人是后来的日耳曼人,斯拉夫人,波罗的人,克尔特人的主要祖先(当然这些民族还有少部分其他来源)。这些生活在欧洲北部的人群是相对比较纯的欧亚部落后裔(主要是中亚部落,少数属于东亚部落),他们被称为诺迪克人种(又称北欧人种,白海-波罗的人种),属于显白人种,也是狭义上的白种人。他们和南欧的地中海人属于不同的人种,在混血程度不高的史前,相比于纯种的地中海人和阿拉伯人,纯种的诺迪克—雅利安人和东亚的纯种黄种人血缘关系要更接近。这一点非常重要,也是下文讨论的基础。北欧的诺迪克人种和南欧的的地中海人种在欧洲中部地区发生混血,形成了阿尔卑斯人种。一般认为,诺迪克人是典型的白种人,而阿尔卑斯人是典型的欧洲人,地中海人则接近中东人。亚洲黄种人和亚洲棕种人矮黑人大约在10万年前就已经分离,而亚洲黄种人和北欧白种人大约在4万年前的伊朗高原上分离,而传统认为东亚的黄种人和欧亚大陆北部的黄种人大约在3.5万年在阿富汗分离,不过最新的研究显示这两者的实际分离时间可能比原来的认识大大缩短,并且分离地点很可能在云南一带。另外需要说明,我国人也有部分芬兰-乌拉尔黄种人的血统,不过和欧洲黄种人属于不同的子类型。这些也非常的重要。回到中国,我们简要得概括一下中国人大体上的血缘构成:
其实,中国人的父系祖先还是比较高大强壮的,只是不如白种人(指北欧人种)高大。在生物学中有两个个规律,一个是海拔越高的地方进化成长的物种就越高大粗壮,反之亦然。还有一个是温带地区形成的人种最高大,热带和寒带形成的人种都比较矮小。在人种形成的初期,受外界的自然环境选择作用是很大的。中国人的祖先出非洲后,曾经在伊朗阿富汗一带的中纬度的高原上进化了很长的时间,然后才通过印度缅甸进入云南的中低纬度的高原,又进化了很长的时间,最后才扩散到中国各地,取代了原来生活在中国境内的古亚洲男人。其中华夏族又在西部甘肃的中纬度的高原上生活了很长的时间。北欧人和印第安人的共同祖先和中国人的祖先在伊朗高原上分离以后,进入了中亚的中高纬度的高原地区进化了很长时间。北欧人印度雅利安人的共同祖先接着和印第安人的祖先分离,并分两批向西进入欧洲或印度。第一批进入欧洲的是西欧的克罗马农人,第二批是近两万年后才进入欧洲的从南俄草原进入的作为征服者的印欧人。
印欧人的另一部分向南进入印度,伊朗,阿富汗,他们就是大名鼎鼎的雅利安人。在两批进入欧洲的北欧人之间,隔着一批在农业扩张时代进入欧洲的中东农民,他们带来了欧洲人中的地中海人种的基因,甚至一部分非洲黑人基因(不过和班图黑人属于不同的子类型)。另需提醒,欧洲的纬度高于东亚。史前的欧洲不仅纬度高,而且经历了寒冷的冰川期,地中海人种的某些体质特征的形成某种程度上就拜这次冰川期所赐,克罗马农人也受到过一点影响。地中海人由于生活在炎热的中东时间更长,所以高鼻深目的特征就更突出。印欧人6000年前征服了欧洲后,由于和被征服者克罗马农人,中东农人混血,加上相同地区的相同地理气候条件下产生的趋同进化,使得所有的欧洲人面目特征都看上去比较相似。
在中世纪,欧洲的气候仍然很寒冷,一直到近代大西洋暖流的形成。北欧人虽然长期生活在北欧寒冷的高纬度平原地区,但是由于他们的种族特征主要形成于温带的高原地区,所以并不矮小。中国人的父系祖先也是在高原上进化出的人种,因此先天的身材并不矮小干瘦,只是纬度没有北欧人高,气候条件也比北欧人暖和一些,所以先天基础没有北欧人高大强壮。饮食习惯和营养条件对人种的形成也非常重要。不过因为和身材矮小瘦弱的古亚洲女人混合,导致了后代的中国人身材,体格被大大拉低。但影响身材高大强壮与否的除了人种的先天基础外,还与人种大致形成后的气候条件,饮食习惯和营养条件息息相关。古代北方中国人长期是世界上最高大的人种之一,不亚于欧洲人。倒是欧洲人历史上有一些时期由于营养不佳,身材比今天矮小很多。
再说一下“高鼻深目”的特征。相比于现代东方人的扁平的五官,纯种的黄种人其实是可以称得上“高鼻深目”的,只是没有中东和南欧的地中海人种那么得夸张。“高鼻深目”本来就是M89的中东部落后代的特征(应该来自于非洲祖先),只是地中海—高加索人种实在是进化得有些夸张了。而M9的欧亚部落后代则要柔和很多,这应该和欧亚部落在较高纬度的地区进化过一段时间有关。不过和大饼脸的古亚洲人相比,纯种的黄种人具有明显的欧亚黄白人种的特征,五官立体感要强很多。今天的亚洲基本已经没有纯种的黄种人了,不过这可以参考一下黄种血统比较多的某些东南亚人,以及介于黄白之间的某些北美印第安人。某些东南亚民族,他们是比南方汉族更纯正的黄种人(当然也是和矮黑人棕种人混血过,只是比例要稍低一些)。
而南方汉族,是比北方汉族更纯正的黄种人。汉人中黄种血统最少,矮黑人棕种人成分最多的是土生的西北汉族(新疆汉族属于中原移民,另当别论)。另外,南方的四川地区有些特别,矮黑血统比其他南方人要多,这是因为四川是属于矮黑种的历史上的氐人的老家。韩国人类似于西北汉族。而历史上的各种北方胡人(白胡除外,包括满蒙),日本人,基本上不属于严格意义上的黄种人。即使今天的两广人,也不是纯正的黄种人,只是黄种成分比其他中国人更多。很多人污蔑广东人是“马来人”,殊不知,广东人的某些“马来特征”,恰恰是黄种祖先的原始相貌特征的一些反映。
二、走向远东的远古人类
【肤色与人种的起源】
现代人(Modern Human) 的学名叫做晚期智人(Homo sapiens sapiens),其生物学级别被假定为一个亚种(subspecies),与各早期智人的亚种同属于一个物种(species)。可能这种级别的设定完全不正确,但至少晚期智人的级别与早期智人的各分支相并列是相当可信的。所以现代人有相当近的同源性,都可以追溯到晚期智人的发源地——非洲古陆。而后在扩散迁徙过程中形成了各分支,我们姑且称其为地理种,即比亚种更小一级的分类单位。不管哪个地理种,与当地出现过的早期智人亚种都没有直系亲缘关系。
现存的现代人地理种有4 个:即黑色人种(尼格罗人种Negroid)、棕色人种(澳大利亚人种Australoid)、黄色人种(蒙古人种Mongoloid)和白色人种(高加索人种Caucasoid)。虽然肤色是个非常明显的特征,却不是关键的判别特征,同一地理种内的各群体肤色会相差很大,造成有些黑色人种群体比一些白色人种群体还白得多。例如埃塞俄比亚(Ethiopia)的一些黑人群体体色非常白。
我们对人种的判定一直采用一种综合的体征标准,以达到确实的亲缘关系。这种标准包括颅面骨骼的各部分形态、发质发色、虹膜色泽,以及其它一些遗传体征。单一的特征都很难判定人种,因为其变异和交流可以造成不同原因的相同表现。比如肤色可能受高紫外线地区的生活环境选择而造成不同的人种都可以有深色的相同表现,不能以肤色深浅判断关系远近。
所以所有体征中,颅面形态相对来说是最好的分种依据,因为其受环境影响比较小,变化范围大,变化以后不易回复祖先群体的形态。一般来说,黑人整个头颅较圆,额部较突;白人较侧扁,面部凹凸不平,眶骨深,鼻根低;黄种人前后较扁,面部较平整,鼻根高;棕色人种颅型略偏圆柱形,面部起伏也较大。另外,从发型上看,黄种人多直发,棕种人多窄波发,黑种人多旋发,白种人多宽波发。这些特征一直被体质人类学界用以分析人种。特别是在东亚的人类学研究中,分析棕色人种和黄色人种的交流历史。
体质形态的分类主导地位在近二十年内渐渐被单倍遗传系统所取代。所谓单倍遗传系统是指纯由父母之中的唯一一方遗传延续的遗传分子。包括母系遗传的线粒体和父系遗传的Y 染色体非重组区。这些单倍分子不受混血影响,其中的无功能区又自由忠实地记录了群体分离的历史。以此判定人种和次人种,层次结构相当清晰。 这些人种的分布在地理大发现时代和殖民洪潮之前是相对隔离的。
具体分布大致是这样的:黑种人分布于非洲大陆和撒哈拉(Sahara)沙漠以南地区;白种人分布于欧洲、北非、西亚乃至印度;棕种人分布于澳大利亚大陆、新几内亚(New Guinea)、美拉尼西亚(Melanesia)、安达曼群岛(Andaman Islands)等地;黄种人分布于其余除南极洲以外的地区,包括北亚、中亚、北欧、东亚、东南亚、太平洋诸岛、新西兰(NewZealand)、印度洋诸岛、马达加斯加(Madagascar)和美洲。但在有些地区,可以明显看到人种间普遍的混合特征,比如中亚和北欧,基本上是黄色和白色人种的混合。这种人种混合的体质观察现象由历史记载也可以得到印证。
人种混合的存在提醒了研究远东地区的人类学家。在远东,特别是东南亚,棕色人种和黄色人种错综而居,有些群体更似乎兼有两方的特征,很难判定。这种格局究竟是不是混血,还是其它机制形成的。这个问题引起了人类学界的长期争议。其中的观点基本上可以分三类:气候带说、进化滞留说和南北混合说。
气候带说认为,早期的人类分散到各地,长期生活在固定的气候环境中,经过千万年的适应,渐渐形成了不同的体质。这种体质的差异从热带到寒带形成了一个差异梯度,越是往热带越具有棕色人种的特征,越往寒带越具有黄色人种特征。较近的历史时期内的移民活动又局部地打乱了这种南北梯度。气候带说的观点在遗传证据发现之前似乎能解释大部分现象。除了个别案例,比如寒冷的塔斯马尼亚,棕色人种定居历史也相当久远,却没有所谓黄色特征。
后来的遗传发现证实了气候带说的观点实为本末倒置。他们所依据的一些特征的确是由气候造成的,却不是长期适应的结果,而是快速选择的结果。比如宽鼻厚唇的热带特征,实际上是由于热带疟166 Journal of National Dr. Sun Yat-sen Memorial Hall疾高发,而地中海贫血基因可以抵抗疟疾。在强大的生存压力选择下,只要经过数代人的强化和固定,任何群体中的地中海贫血基因都会极高频地出现。而地中海贫血患者的胎儿在发育期间会因为缺氧而造成宽鼻厚唇等特征。这些特征与群体久远的发生历史毫无关系,根本不能解释黄种人和棕种人形成的机理。
进化滞留说与结构主义人类学有关系,支持者主要是一些古人类学家。他们认为远东人类由当地的古人类进化而来,其中棕色人种的特征与古人类更接近。他们发现,十万年以来,远东地区的人类化石“渐渐”由棕色人种特征转化为黄色人种特征。旧石器时代基本上“原始”特征占大部分,到新石器时代,“原始”特征个体的比例越来越少。
华夏中原地区到夏商时期还有“原始”特征的类澳大利亚个体存在,到后来渐渐消失。这体现了人类的一个进化过程,一些“原始”特征渐渐在群体中被淘汰。而在澳大利亚和伊利安(Irian),“原始”特征却滞留在现代人群中。令人匪夷所思的是,各个人种的进化速度怎么会差别如此大。而各地黄种人却几乎匀速同向地进化了。这种现象的原因根本无法解释,而基因时代是拒绝这样不能解释的理论的。当然,现在的进化研究已经证实了东亚的古人类和现代人无关系,现代人的差异也不可能是因为进化程度的不同造成的。
南北混合说是早期群体遗传学界的普遍观点。他们在大量的双倍遗传标记研究中发现,南北差异始终是主流。对这种现象成因的最自然解释就是南北的人群有完全不同的来源,黄种人来自北方而棕种人来自南方,渐渐地扩散融合形成现代的格局。最早的看法以为现代人是当地起源的,所以猜想黄种人来自北京猿人,棕种人来自爪哇猿人Java man)。现代人非洲起源说成为共识以后,又猜想黄种人从北方中亚进入,而棕种人从南方东南亚进入。但是从最合适的单倍遗传标记分析看,东亚人群几乎全部是从南方进入而后向北扩散的,所以南北异源之说也无从说起。这种观点基本上是种形而上学的观点。
从单倍遗传标记的分布看,对远东两个人种的分布格局形成机理,可以再提出一种新假说——先后替代说。棕色人种最早到远东,扩散开来;黄色人种来到远东以后渐渐取代棕色人种,形成黄色人种占据主体地区,棕色人种处于偏远地区的格局。
【远东人群遗传层次结构的发现】
自从单倍遗传系统的研究建立起来以后,学界对全世界的各个群体作了深入详细的调查分析,数十万的数据呈现在了我们的面前。从这种全面的数据中探索规律,就更能够避免管窥蠡测断章取义了。我们先来看看Y染色体非重组区的单倍群分布, 在全世界的分布具有非常明显的地域特异性。
Y染色体主干单倍群在各地人群中的分布比例
非洲的E型,美洲的Q型,欧洲的R型,中东的J型都是非常有特色的。这些特色单倍群说明了该区域的人群的主体起源是一致的。更深入地说,在这一地区某种来源的人群的后代已经完全占据了主导地位,其它成分或许曾经是多数,但现在已经被覆盖(注:即过去占优势的人种被后来的另一人种基本上消灭掉)。
在非洲,这表现为E对A和B的覆盖;在欧洲则表现为R 对I 的覆盖;而在美洲,基本上只能看到Q,其它类群很难观察到。所以在人类定居历史悠久的大陆,一群人扩张开来,替代另一群人的过程往往会发生。这既体现在现代人种对古代人种的替代上,也体现在现代人种的历史中不同人群的更替。族群发生、发展、兴盛最后衰亡的历史在不断的演绎着。
Y 染色体主干单倍群的区块分布示意图
在远东地区,这种更替的迹象更为明显。我们可以看到在普遍存在的C 和D 中间,O 型和P 型硬生生地插了进来。这种更替的过程并没有完成,随着文明社会的到来,这个过程被定格、凝固了起来,使我们看到今天的格局。
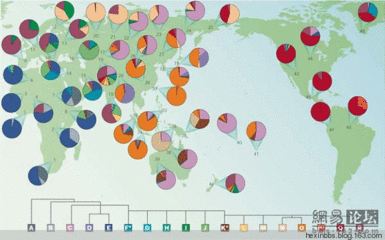
C 型主要分布于澳大利亚、伊利安、太平洋、西伯利亚;D 型主要分布于青藏高原、日本、安达曼群岛;O 型主要分布于东亚和东南亚;P 型主要在蒙古高原和中亚草原,并低频存在于O 型群体中。CD(注:即棕色人中基因类型) 和OP(注:即黄色人种基因类型) 的对立就揭示了远东地区的现代人发展历史中最主要的一个更替过程,这个过程就是我们这里要建立的假说,新石器的黄种人取代了旧石器的棕种人。
把单倍群的分布区块化地画在地图上,我们更容易看到,C型处于远东的南北两端,而中间地区,实际上也已非常低的频率存在着。D 型则处于东西两端,同样在中部低频存在。对这种现象,可以有两种解释。其一,人群进入东亚以后,内部共有着C D O P等类型,当进一步向周边扩张的时候,由于迁徙的人口较少,正好只有一小部分的CD 个体来到了周边,发展成今天的样式。这就是一种瓶颈效应的解释。其二,CD 的群体先来到远东地区,并且定居到了各个小区域,OP 到达以后渐渐从中间向周边取代CD。这是群体更替的解释。对这两种解释的取舍,仅仅从群体类型的格局上是无法判断的,还必须借助于其它的证据。从Y 染色体的STR 标记,我们可以计算出各个单倍群的发生年代 。 把现在得到的所有有关个体都集中起来168 Journal of National Dr. Sun Yat-sen Memorial Hall计算,我们得到C D 的总年代为约95000 年,而O P 的总年代却是约53000年。如果是瓶颈效应产生的格局,那么CD 和OP 的年龄应该是一样的,因为进入时他们是一个群体。而取代说才会是有不同的年代,CD 的年龄要比OP 大得多。所以,这基本上可以肯定是群体更替造成的,是OP 正在替代CD。
我们把CD 确定为棕色人种类型,把OP 确定为黄色人种类型,是通过这两个人种的大量的群体分析得出的。在典型的棕色人种中,澳大利亚、伊利安和美拉尼西亚,最主要的类型是C,安达曼几乎全部是D。而黄色人种毋庸置疑,O 和P 始终是主流。所以OP 替代CD 就可以解释为使黄种人替代棕种人。
在母系的线粒体的分布中我们不仅能看到替代,也能看到交流。人类历史上,基本是母系的交流要多于父系的交流。就替代而言,同样的,线粒体单倍群M*和N*被ABCDFG等挤压在周边。在东亚大陆上,主要的是B,F,G,D,M7,M8。由于交流的大量存在,替代的格局不如Y 染色体明显。
群体更替的观点确立以后,还有一点需要深入分析。因为更替也有两种可能,一种是外来的更替,另一种是内源的更替,两者都会得到替代者比被替代者年轻的格局。因此,必须把几个人种放在一起分析,看看黄种人是在棕种人内部还是在外部。白种人和黑种人必然在远东人类活动的大规模影响之外,如果黄种人的替代是内源性的,也就是从早期的棕色人种内的一群爆发开来的,那么他势必距离棕种人更近一些,而距离其它人种更远一些。反之,则会距离棕色人种远一些,至少不会近。
所幸我们现在有足够的材料来做这样的分析。当我们用单倍遗传标记把各地区的人种进行聚类分析,得到了非常有趣的构型。非洲黑人自然很早就开始分化成几个细支,并不断地进行着基因交流。走出非洲的群体中,棕色人种最先分化,远远地离开了白色和黄色人种。白色和黄色的分化则相当晚,几乎是与棕色人种分化年代的一半。问题已经相当清楚了,在远东替代棕色人种的黄色人种是从外部进入的,入口就是东南亚。
这种替代的过程是怎么进行的呢?一种可能是黄种人进来以后,与棕种人通婚交流,渐渐地由于遗传机理而占到了优势。另一种可能是黄种人把遭遇到的棕种人消灭,占据他们到达的地方。或者,可能是一种中间的形式,消灭和交流兼有。从Y染色体看,主要地区的棕色类型CD 都非常少,除了波利尼西亚(Polynesia)的黄色人种中间,看似棕色的男性可能大部分被消灭了。从线粒体看,人种之间的差异也是相当大的,但是交流的迹象相对多得多。所以女性可能被保留得稍多一点。要进一步看这种交流的程度,还是要看双倍遗传的常染色体基因。我们来看一下远东特色的HgC 基因的几种类型在远东各地的分布。
其中,1 型在东亚大陆,2 型在太平洋岛屿,3 型在北亚,这些都属于黄种人。
*型是根部的未区分类型,澳大利亚棕色人种中全部都是这一类,南亚和东南亚也不少,但是在其它地区则基本上没有出现。为了分清这四个亚型之间的关系,我们进一步观察它们的网络关系图。
由这个网络图观察,我们要推翻前面的1,2,3 型属于黄种人的论点。在 H g C 起源位置上的原始类型都存在于南亚和澳大利亚的棕色人种群体中,而黄种人的几种亚型都起源于棕色人种,另外黄种人也没有原始亚型。这说明,H g C 不是黄种人原有的基因;黄种人在与棕色人种接触的早期,交流得到了少量H g C;黄种人的进一步扩张过程中,人口膨胀使H g C基因突变成新的亚型,但是并没有与棕色人种进一步交流,所以没有原始类型。现在可以得出结论,黄种人进入远东时曾与棕色人种有过有限的交流,而扩张期间则基本上把遭遇的棕色人种消灭了(所以说原始人是野蛮的原始时代生存竞争是十分残酷的)。
现在我们可以勾画棕色人种和黄色人种在远东地区的迁徙历史了。
A. 棕色人种大约十万年前走出非洲,至少在五万年之前已经来到东南亚;黄色人种则是大约五万年前的时候开始向东迁徙;
B. 大约四万年前,棕色人种已散布于东北亚到澳洲的广大地区,而黄色人种则进入了东南亚;
C. 大约三万年前,黄色人种在东南亚、印度尼西亚次大陆和中国南方进一步扩张,而棕色人种可能有一部分已经来到美洲;
D. 大约两万年前,黄色人种几乎征服了东亚和东南亚的大部分地区,使该地区的棕色人种锐减;
E. 大约一万年前,黄种人进入美洲,并成为美洲的主宰;
F. 在数千年内,黄种人向中亚、北欧、印度洋和太平洋诸岛扩张,东亚最偏远地区的棕色人种也渐渐与黄种人融合。
远东地区两个人种扩张和替代的过程示意图
——A约50000年前;B约40000年前;C约30000年前;D约20000 年前;E约10000年前;F约500年前。
这是一个动态的替代过程,黄种人扩张的同时棕种人也在继续前进。替代过程中的征服和杀戮恐怕是难免的。这与两个人种社会文化上的严重差异有关。黄种人之所以能替代棕色人种,也可能与其社会文化的相对先进和工具的相对先进有关。直到殖民时代到来,大部分的棕色人种始终都使用粗糙的旧石器,没有陶器;而黄种人则很早就有陶器和新石器。可能东亚地区的旧石器时代没有后续的晚期智人考古文化大多属于棕色人种,而新石器及其祖先型的旧石器文化则可能都属于黄色人种。社会和器物的更替通过人种的更替实现了。
【棕色人种的扩张史推测】
棕色人种早期详细的迁徙历史已很难考证,他们是进入远东最有可能的线路是沿着阿拉伯海的沿岸。早期的棕色人种应该是以采集渔猎为生的,而且最可能是沿海打鱼为生。这是一种比较容易的谋生手段,海岸滩涂总是可以找到一些高蛋白的食物。一方面他们因为沿海觅食而渐渐向东迁徙,散布到了非常广阔的地域;另一方面,滩涂海产也是有限的,人口不可能过度增长。他们最大的分布特征是大范围低密度,这严重限制了他们的生产力发展。因为他们没有生存压力,就没有生产革新的动力。
人口稀少,新的发明不易流传,很容易埋没。他们就是这样使用着最简陋的工具探索东方的新世界。这也是他们后来大部分被淘汰的原因之一。在沿海捕捞的过程中,什么时候他们学会了使用木筏不得而知了。但可以肯定的事,木筏帮助他们进一步扩展领地,在从印度尼西亚半岛到伊利安-澳大利亚大陆的迁徙中起了很大的作用。如果他们到达了美洲,那么这也应该完全是木筏的功劳。因为那时的白令陆桥一片冰雪覆盖,原始社会的人类根本不能通过。如果只是坐着木筏沿海捕捞为生,情况就不同了。所以棕色人种在三万年前到达美洲是完全可能的。
我们可以想象,最早的棕色人种拿着最简陋的旧石器和尖木叉,在印度洋岸边走着,寻觅着鱼虾蚌蛤。经过了一代又一代人的扩展,几万年后他们来到了南亚和东南亚。在海岸边,到处是一个个棕色人种的小村落。当时的海平面比现在要低好多,巺他(Sunda)群岛与印度支那半岛连成一片,成为一个很大的印度尼西亚半岛。澳大利亚和伊利安以及美拉尼西亚的许多岛屿也连在一起。棕色人种很快就散布到了这片地区。到海平面抬升以后,印度尼西亚半岛消失,澳大利亚和伊利安远离亚洲大陆,棕色东南亚的尼格利陀(Negrito)小黑人人种在澳洲这两块世外桃源般的大陆上居住到了现在。而印度尼西亚和菲律宾(Phillipines)的偏远山区也还有零零落落的棕色人种部落,他们被称为尼格利陀人(Negrito)。
在南亚地区,印度半岛上的居民达罗毗荼人(Dravidian)长期以来都是棕色人种,直到白种的雅利安人(Arian)入侵才使他们发生了混血。他们是所知的棕色人种中文化发展的最为先进的一支。缅甸以西的海中,棕色人种也留存在安达曼群岛上,并保持着非常纯的血统。实际上来到远东以后,一些部落必然发展出以狩猎为生的生活方式,因而进入大陆腹地。他们来到荒无人迹的地方,没有任何的竞争者。只要用简陋的工具就能获得维持生存的食物。
棕色人种的食物必然不会非常充足,没有精湛的狩猎技术无法保证每一次都有收获,所以他们不浪费任何食物来源。从安达曼到伊利安,棕色人种始终保持着食用人类尸体的传统,并以此建立他们的道德标准。尽管到处流传的食人族的传说总是让人不寒而栗,但对于棕色人种来说这是自古流传下来的部落生存必需的准则。这也成为后来他们为黄种人所不容的原因之一。
棕色人种北上东亚大陆的历史可能比南下澳州要晚一些。来到澳洲可能是六万年前,而在东亚大陆散布开来恐怕是四万多年前的事。中国的旧石器时代晚期文化或许大多是棕色人种的,虽然他们肤色可能并不深,与南方的棕色人种在体质上也可能有较大差异。至少,他们应该看上去不太像黄种人,面部的凹凸感较强,头颅形态也不同于黄种人。
我们这里讨论的始终是晚期智人,与早期智人无涉。到了今天,早期智人的来源成了古人类研究中的最大的疑点。直立人起源于非洲,晚期智人也起源于非洲,而早期智人是直立人进化而来的还是另有渊源却是个未解之谜。但至少欧洲的早期智人源于非洲。
华夏地区的旧石器时代晚期人类遗骸有柳江人、山顶洞人、资阳人、札莱诺尔人等,他们的形态差异颇大。颜誾的观点比较客观,他指出柳江人比其它人更接近黄色人种。山顶洞人和资阳人很相似,有观点认为他们相对最接近虾夷人(Ainu)。我比较倾向于这种观点,这说明在三万年前,黄种人已经到达了中国南方珠江流域,而棕色人种还在长江流域和黄河流域活动。当时的日本列岛还与朝鲜半岛连在一起,是人类最容易到达的地区。捕捞生活的棕色人种就这样沿着海岸向东北延伸,通过白令海湾顺势到了美洲。
棕色人种到达美洲目前还没有遗传证据,但是许多古人类遗骨和考古遗迹的发现指向这一论点。在亚马逊(Amazon)发现的早期人类遗骸与印第安人完全不同,而更接近于澳大利亚土著。下加利福尼亚的一个特殊部落的发现更揭示了一个体质、文化和传统完全不同于印第安人的人群的生活。据说这个类似澳大利亚人的部落被西班牙人带来的瘟疫灭绝了。棕色人种在三万年前到了美洲之后过了两万年,黄种人才开始到来。黄种人和棕种人对美洲的争夺战可能在美洲大陆广泛地开展着。中美洲古老的奥梅克(Olmec)文化留下了许多石刻头像,无论如何都没办法把他们与黄种人联系起来。如果棕色人种来过美洲,他们一定就是。
【黄色人种的扩张史推测】
黄种人走向远东的路线应该与棕种人不同,他们更可能采用了一条陆路。从西亚,进入印度河流域,在南亚次大陆的北端经过,进入东南亚。初来乍到的黄种人还没有能力与棕色人种抗争。至少在南亚的时候,印度半岛南部生活着大量的棕种人,一小批的黄种人只能沿喜马拉雅山南麓匆匆通过。这期间也融入了少量棕色人种的基因。在东南亚的密林里,黄种人发展壮大了起来,渐渐占据了优势。也是在这一时期,黄种人分化成了沿海支和内陆支。
沿海支的黄种人最初在越南、广东一带形成,后来部分人南下马来和印度尼西亚,部分人则沿着东海大陆架北上。这些人(水注:沿海支)就是南岛语系(Austronesian)和侗台语系(Daic)的人群的祖先,就是民族学上所说的百越民族。这一支的人群携带着明显的Y 染色体单倍群O1。内陆支的黄种人则形成于缅甸、云南一带,并长期居住于此。由于历史比沿海支稍长,渐渐形成了更多的基因突变。这群人(水注:内陆支)是南亚语系(Austro-Asiatic)的祖先,就是民族学上说的百濮民族。他们带的Y 染色体特色单倍群为O2 和O3。
我们这里提到某个人群带有某种特色单倍群时,并不是说他们只有这种单倍群,而是说他们以此为主,还兼有其它类型。在这些群体中,的确是特色单倍群占到绝大多数。
当时的末次冰期不仅仅造成了海平面的下降,大陆边缘的延伸,另一个效应是当时的一些不高的山都是积雪冰封。所以长江流域和珠江流域之间的南岭居然成了一个不小的阻隔。这限制了黄种人向北的迁徙。当然,可能还是有一些人进入了北方,这些人可能携带了特色Y 单倍群P。他们(注:在冰期末最先翻过冰雪进入中国北方的黄种人)与后来到来的黄种人混合以后形成了黄种人的北亚支,就是阿尔泰语系(Altaic)的祖先。
末次冰期结束使黄种人大量北上,这个时间在大约两万年到一万年前。我们可以看到北方的人种发生了明显的变化,半坡和宝鸡的新石器人种与甘肃史前古人种完全不同。黄种人的史前大北伐使中国大陆的人类历史发生了革命性的变化。不久,新石器产生了,并迅速传遍了各地的黄种人社群,农业文明的星星之火在各地的黄种人中燃烧了起来。这开创了一个全新的时代,社会发展的火车头被拉动了。
黄种人的北伐也是分两条路线进行的。东部的沿海支进入浙江、江苏、山东,形成了河姆渡—崧泽—良渚的先越文化和大汶口—龙山的先夷文化。这一支人群与南下的沿海支黄种人遗传上比较接近,所以古人类学家观察到大汶口文化的人种体质上与玻利尼西亚的南岛民族极为相似。夷越类群(注:沿海支北上,商人与此类群关系密切)在沿海一带的发展相当快,东北渤海周边的红山文化也可能与这一类群有关。农业、新石器、玉器和早期的中国哲学思想都可能先产生于这一族群中。
西部内陆支的先濮民族也几乎同时北上,他们是通过云南、贵州、四川这条路线北上的,并且至少分出了两支。一支沿着长江往下走,到了三峡地区和洞庭湖平原, 发展出了城背溪— 大溪— 青龙泉的先蛮文化, 形成以后的苗瑶民族(Hmong-Mien)。另一支(注:内陆支北上,和夏、周有关)沿着四川的几条大河北上,来到了黄河上游地区,发展出了半坡—仰韶的先羌文化,形成了以后的羌华民族(Sino-Tibetan)。我们计算北方的O型的年代时,基本上都得到一万年左右的数值。
北部的阿尔泰群与当时北上的两支黄种人广泛地发生了混血。至今可以明显地看到,东部混有较多的沿海支成分O1,中部混有较多的内陆支成分O3。在混血发生之前从北部支已经有一部分的人开始通过白令路桥向美洲跋涉,这部分人在行进过程中Y 染色体从P 变成了Q。一万多年前到了美洲以后,他们(大陆北部黄种人)迅速地向南扩张,在千年时间内遍及了南北美洲。
从Y 染色体看黄种人的迁徙,Underhill 画出了一系列的线路图。他并没有在路线的细节上画得很详细,因为有些细节还不够清楚。比如P 北上的路线,究竟是波斯还是中国,当时还不很清楚。现在在中国南方发现很低频的P 的原始类型,说明P 更可能是通过中国大陆北上的。
黄种人来到北方以后,对当地的棕种人进行了杀戮和驱逐。但是这个过程并不是很快就完成的。棕种人已经在北方生活了这么长时间,并继续在许多偏远的山区躲藏了很长时间。直到商代的人骨材料中,我们还可以发现可能是棕色人种的成分,而且为数不少。只是他们依然是黄种人杀戮的对象。在殷墟的祭祀坑里有大量的异族俘虏的头骨,其中很大一部分可能是棕色人种。
这个替换的过程在中国进行到了非常晚的时期。唐代昆仑奴的传说不知道是否与此有关。据地方史料记载,清代广东官宦家庭中还有过豢养“小黑人”的案例。在台湾,最早到达的南岛语系原住民有着灭绝“矮黑人”的传说。这些零零星星的记忆都指向一点,黄种人对棕种人进行了种族灭绝。所幸的是,在菲律宾和印度尼西亚,这个棕色人种的分支——“矮黑人”还是存在的。我们无需对此忏悔,这是那个社会阶段必然要发生的事件。
在原始社会,与本种群有竞争关系的其它种群就是敌人,只有你死我活的结局。要么黄种人消灭棕种人,要么棕种人吃光黄种人。在这个长期的拉锯战中没有其它的结局。而当黄种人文化上先进于棕种人的时候,结局就只有一个了。那就是黄种人成了远东唯一的主宰。 当历史进入了阶级社会,情况就发生了变化,奴隶的需求使得黄种人对遭遇到的棕种人不再灭杀。那段时期黄种人才到达的边缘地区,比如青藏高原、日本列岛,棕色人种的成分被大量地保留了下来。而南岛语系的族群向两个大洋拓展的过程中,甚至避开了棕色人种居住的区域。
【两个人种的交融地区】
日本列岛在棕色人种到来时还可以通过大陆架很容易地到达,而当黄色人种来到东北亚时,已成为茫茫大洋相隔的海岛,是黄种人传说中的蓬莱仙岛。所以长期以来,岛上居住的绳文人一直是与大陆不同的人种。
关于绳文人的来源,日本研究者的大胆猜想很多。有一点是公认的,现代北海道的虾夷人和种子岛的部分居民应该是绳文人的后代。那么根据虾夷人的遗传结构,我们能够了解绳文人的来源。很幸运,虾夷人的Y 染色体给出了明确的结论,他们大多是D,还有部分的C。他们必然是早期的棕色人种的后代。
日本的黄种人——弥生人,是中国的春秋战国时期迁入的,进入口是九州岛岛。他们向本州岛及更东北方向扩张的过程中,对旧石器的绳文人进行了征服和奴役。势必造成了绳文人比例的下降。但是到1800年,我们可以看到本州岛岛的北部开始到库页岛,都还是虾夷人的领地。
从日本各地的遗传结构看,棕色人种的遗传比例虽然是从西往东递减,但各地的该种成分都不在少数。所以日本人总体上应该算是两个人种的混血区域。
虾夷人(Ainu)及其1800年分布区
对于虾夷人奇特体质,以前的人种学家猜想很多,最后有人认为这是一个处于黄种人和白种人之间的过渡种群。这实在是一种匪夷所思的观点,他们居然与远在地球另一面的白种人有关。其实他们的体质形态独特性说明,棕色人种经过漫长的历史发展,已然形成了几种不同的体质,至少澳洲土著、尼格利陀人和虾夷人是有区别的。
青藏高原的情况要复杂得多。藏族是一个很晚才形成的民族。他们是秦汉时期开始向高原迁徙的羌族中的两支——发羌(黄色人种)和唐旄(棕色人种)。在进入高原后,与当地的雅砻土著发生了混血。在藏族起源的神话传说中,雅砻土著被转化成了丑陋的猕猴,羌人则是仙女,两者婚配繁衍出了藏族。其实雅砻土著属于棕色人种,在羌人的审美观念中,他们的确是非常丑陋的。他们的混血使羌人很快适应了高原的环境。也使青藏高原的历史进入了一个全新的时期。我们在研究现在的藏族人的遗传结构时,还是能看到大量的D 型Y 染色体。而观察藏族的面貌特征,甚至在同一个村子中也会看到两种截然不同的样式。有的人是挺拔的羌人的面容,有的人则是敦圆的棕色人种面貌,肤色的差异也很大。
当我们在青藏高原进一步扩大调查范围,研究氐羌的其它民族,更有趣的现象出现了。曾经在中国南北朝的历史舞台上叱咤风云的氐族,现在仅存于四川和甘肃边界地区,有着惊人的Y 染色体遗传结构——全部的D 型。难道他们也是棕色土著的后代?
我们恍然大悟,原来在汉藏语中,“氐”这个词,从来就是“底层”的意思。羌人来到青藏高原,把原本就在这里的土著居民称为“氐族”,因为他们“原抵”就在那里。我们再看羌族的结构,他们也含有部分的D 型,但还是以O2 为主。再看华族,就几乎不含有D 型了。所以,“氐—羌—华”原初大概就是这么分化的。
而历史上对西戎民族的认识,一开始就有氐和羌的不同概念。我们进一步计算当地的D 和O 的年代,发现D 至少在四万年以上,而O 却最多纔有一万年历史。青藏高原上的D 型成分与安达曼群岛棕色人种的D 型应该是有关系的。
氐羌民族的几种面貌——A 白马氐族妇女(摘自注23);B 羌族妇女;C 黄种特征康巴男子;D 棕种特征的康巴妇女;E 牦牛羌纳西族妇女
在东南亚的中南半岛,这样的遗传成分也是存在的,并且在各个民族中的成分比例不同。东南亚是世界上民族结构最复杂的地区之一,南亚语系的百濮民族是最古老的黄种居民。由于生活于热带,他们的肤色很深,比如佤族就是我国肤色最深的一个民族。但是有些民族的肤色就深得过份了,比如泰国和马来西亚边境上的土著Senoi 族,再加上他们其它一些特别体征,不得不让人怀疑他们是否含有大量棕色成分。可惜他们的遗传结构至今还没有调查过。
[