现代人类学通讯
第 14 卷第 S 期
第二卷
Chin. J. Healthy Birth & Child Care 14(S):72-91 2008 年 Communication on Contemporary Anthropology
COM. on C. A. 2:e11, 2008
Vol. 2
动态评论
Y染色体单倍群D在东亚的分布及其意义
中国语言论坛, 苏州 215000
评论文献: H, Zhong H, Peng Y, Dong YL, Qi XB, Zhang F, Liu LF, Tan SJ, Ma RL, Xiao CJ, Wells S, Jin L, Su Shi B(2008) Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biol 6:45. Chandrasekar A, Saheb SY, Gangopadyaya P,Gangopadyaya S, Mukherjee A, Basu D, Lakshmi GR, Sahani AK,Das B, Battacharya S, Kumar S, Xaviour D, Sun D, Rao VR(2007) YAP insertion signature in South Asia. Ann Hum Biol 34:582-586. 摘要: 石宏等(2008)近期发表的论文是迄今为止对Y染色体单倍群D描述最详尽的论文.文献得到的单倍群D 下各个支系的分布以及年代有助于我们了解东亚早期现代人的分布和迁徙状况.在西藏人和泰国人中分别发 现了DE* 和D*.这使我们能够在Chandrasekar等(2007)的讨论的基础上就单倍群D的诞生地作进一步的分析. 同时,本文从绝大部分包含东亚YAP和单倍群D样本的论文中收集了东亚各个人群中单倍群D的分布数据.分 析这些数据并参考石宏等(2008)的结论后认为,单倍群D可能诞生在东南亚与印度之间,向东迁徙后不久诞生 了D1,而D*进一步扩散到整个东亚,后来在孤立的情况下分别诞生了D2(日本)和D3(西藏). 关键词:Y染色体; 单倍群D; 东亚; 迁徙; 藏缅; 苗瑶; 董氏族; 多弥; 白兰
Distribution of Y chromosome Haplogroup D in East Asia and its Anthropological Implications
LAN Hai
Linguistic Forum of China, Suzhou 215000 China
ABSTRACT: The recent paper by Shi et al. (2008) provided the most details for the ethnic features of Y chromosome haplogroup D. The geographic distribution and age estimate for haplogroup D and its sub-clades helps in understanding the origin and migration processes of the early modern human in East Asia. Clades DE* and D* were found in Tibetan and Thai respectively. These findings make the search of the birth place of haplogroup D possible. Here we continued the discussion of Chandrasekar et al. (2007). The frequencies of haplogroup D in various populations of East Asia were collected from all the available literature. The analyses showed that the emergence of haplogroup D may be between India and Indochina. Clade D1 emerged subsequently when population move to the east. After clade D* diffused to the whole East Asia, it gave birth to clade D2 in Japan and clade D3 in Tibet independently. Key words: Y chromosome; Haplogroup D; East Asia; Migration; Tibeto-Burman; Hmong-Mien; Ldong proto-Tibetan; Domi; Phrom
本文从绝大部分包含单倍群 D 样本的 论文(包括中文发表的论文[1-69])中收集了 东亚各个人群中 Y 染色体遗传标记 YAP(包 括单倍群 D)的分布数据, YAP 在东亚的 把 各支系在各个人群中分布作了详细列表, 在 前人研究的基础上就 Y 染色体单倍群 D 的 起源和扩散作了一些讨论.并且翻译了与 YAP 相关的两篇文献,第一篇专门描述了 东亚特有的 Y 染色体单倍群 D[65], 第二篇 讨论 YAP 起源于南亚的可能性[57].
一,YAP 的发现以及相关研究
在减数分裂中, 人类Y染色体上长臂和 短臂末端的拟常染色体区会和X染色体发 生重组,而其余的Y染色体特异区
收稿日期:2008 年 12 月 14 日
(Y-specific region)不会发生重组,以单倍型 形式遗传,因此在Y染色体特异区上发生的 突变能够保留下来, 并能按父系遗传方式遗 传给后代.Y染色体上的单核苷酸多态位点 (SNP)组成的遗传标记具有很强的群体特异 性, 这些突变大部分按照一定的时间顺序依 次发生, 能清晰地记录群体的历史, 这些特 点使Y染色体在重建人类的进化史和研究 父系遗传关系方面占有优势[70]. 人类遗传学家翰墨博士(Dr.Hammer)在 文献[71]中最早对YAP进行了描述,它是Y 染色体特有的DYS287 Alu插入单元,位于 人类Y染色体长臂的非重组区(Yq11).之后 YAP这个与父系遗传相关的Y染色体多态 位点被广泛研究 [72,73]. DYS287通常缩写
修回日期:2008 年 12 月 16 日 联系人:兰海 ranhaer@gmail.com
2008 年 12 月 17 日
http://COMonCA.org.cn/Abs/2008/011.htm
72
上海人类学学会 Shanghai Society of Anthropology
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 为YAP,标识为YAP-(祖先型)和YAP+(突变 型). YAP插入事件是Y染色体世系内部结构 上一个古老的分支, 对研究人群的进化非常 有用[74,75].YAP染色体在全世界都有分 布,在非洲[76],西藏[73,77 ]和日本[78]等 地区高频出现.有报道称,非洲和亚洲的 YAP单元在Y染色体上的插入位置是一样 的[74].这说明,非洲和亚洲的YAP属于同 一个突变事件,它们有共同的祖先.YAP 跨洲的两极分布被认为与它的非洲起源相 符, 但是另外一种YAP起源假说也被提出来 了. 在检测了SRY(sex-determining region on Y chromosome)区域的YAP位点和核苷酸位 基于组合单倍型 点4064上G-A的突变之后, 的数据, YAP亚洲起源假说被谨慎地提出了 [73,79].在对1544个YAP和SRY1060样本进 行9个双等位多态位点的分型之后[80],翰 墨博士分析了Y染色体变异的分支结构,认 为这些结果进一步支持以上假说. 这些结果 可以解释为支持走出非洲的大规模扩散, 但 也可以看作是另外一种图景的证据: 一次晚 近的迁徙将部分亚洲Y染色体(YAP+支系) 带回非洲,而没有完全替换掉原有的YAP的非洲Y染色体基因库.这是对人类早期迁 徙历史的一次重构, 鼓励我们继续进行关于 YAP起源地的尚无结果的讨论[81]. 文献[70] 对全球超过1000个样本进行了检测, 由一个 三等位多态位点(即Alu 插入元件)和166个 双等位多态位点建立起116个Y染色体支系 的系统发育结构,进而讨论了现代人Y染色 体多样性. 该文献提出了与YAP亚洲起源假 说不符合的证据. Weale于2003年[82]报道了在尼日利亚 发现了5例属于超单倍群DE*的样本.在仔 细地研究了YAP内部分支的各种可能顺序 之后,Weale认为这样样本更倾向于支持 YAP的非洲起源. 不过翰墨博士与岇得希博 士(Dr. Underhill)在2008年合著的极其重要 的文献[83]中,并没有对YAP的起源作进一 步描述. 日前, 石宏等[65]在西藏发现的2例DE* 为这个争论又增添了疑问. 但由于DE*的样 本数实在是太过稀少, 我们还不能确定它的 起源地, 就像[82]认为的那样. Chandrasekar 的论文[57]讨论了YAP起源于南亚的可能 性. 论点之一是, 西孟加拉邦的拉吉班什人 和奥里萨邦的保里布伊亚人中发现了与安 达曼人的mtDNA拥有最亲缘关系的单倍群 类型, 此发现说明安达曼人的祖先曾在南亚 大陆上停留过一段时间.论点之二是 F-M168在南亚深度分化并仍存在大量的
73
动态评论
F*,而史前向非洲的回迁也有证据支持. 这说明YAP(包括D)很有可能诞生于南亚的 M168.我认为这个推测颇可采信.
二,东亚的单倍群 D
Thangaraj等于2003年初发表了一篇极 其重要的文献[11],报道了安达曼群岛居民 中D*的发现.这是对走出非洲的"南部路 线"的有力支持. 在不久之后发表的文献[84] 中,Thangaraj等对线粒体基因组(包含6500 例印度样本)的研究发现,安达曼人中存在 两种独特而古老的M世系――M31与M32, 从而提示安达曼人的祖先可能直接来源于 "走出非洲"后到达印度洋北岸的现代人类. 不过2006年初发表的文献[85]说明这两个 单倍群并非安达曼人特有的, 中国科学院网 站对这篇文献的报道如下: "由于具有和非洲俾格米人非常相似体 貌特征, 生活在印度安达曼岛上的安达曼人 的起源一直为人类学家们所关注. 印度学者 Thangaraj等对线粒体基因组的研究发现, 安 达曼 人中存 在两 种独特 而古 老的M世 系――M31与M32,从而提示安达曼人的祖 先可能直接来源于"走出非洲"后到达印度 洋北岸的现代人类. 为了进一步探讨安达曼 人的起源, 在昆明动物所所长张亚平院士的 领导下,云南大学印度籍研究人员Malliya gounder Palanichamy博士和昆明动物所 姚永刚, 孔庆鹏博士等对于来自印度各地的 大量人群样本的线粒体DNA多态进行详尽 分析, 在此基础上针对性地选择代表性样品 进行线粒体基因组全序列测定. 他们的研究 结果发现,来自印度东北部Rajbhansi人群 中存在一种古老世系(M31b),该世系与 Thangaraj等报道的M31世系(M31a)共享 有三个保守的编码区突变,从而提示M31 世系的祖先类型更可能起源于印度次大陆. 因此, 安达曼人的祖先可能是来自定居于印 度次大陆并已发生一定分化的人群,而非 Thangaraj等所认为的是直接来自于"走出 非洲"后沿亚洲海岸线迁移的现代人类祖 先." Thangaraj 等 对 此 进 行 了 回 复 [86], 说 M31a虽然不是安达曼独有的,但这个单倍 群与M31b仅共享有三个编码区突变也能说 明安达曼人在极其古老的年代就已经与南 亚大陆隔离了.并坦言,他意识到"找不到 M31出现的证据并非不存在M31的证据". 总的看来, 我认为安达曼人的祖先来到安达 曼群岛之前,必定在大陆上生活过一段时 间;况且,在大陆上(比如毗邻的东南亚)仍
FOCUS
COM. on C. A. 2:e11, 2008
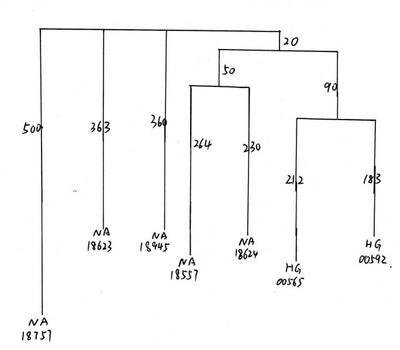
旧有一定比例的D,因此在母系上与大陆人 群存在一定联系,这个结果倒是非常合理 的. 借助于对YAP的深刻理解, 翰墨博士对 日本的YAP进行了详尽的研究, 发表了关于 日本人的双重起源的经典文献[1].但是在 东亚的大陆部分, YAP一直没有得到深入的 研究.YAP(DYS287)被发现之后,从1998 年发表的文献[40]开始,国内也进行了大范 围的采样和研究. 不过, 由于分型技术等原 因,最初的一些文献仅限于对YAP+在各群 体间的比例的讨论. 同时也由于YAP在东亚 人口中的总体比例并不高, 而且分布地域有 限,后来的文献主要关注于其他的单倍群 (比如单倍群O等).关于东亚的YAP和单倍 群D的数据散落在各个文献中[1-69],很多文 献之间相互引用数据, 经整理和分辨, 得到 了表1,其中罗列了国内外文献中东亚各个 人群中YAP各个支系的分布频率.在东亚, 几乎所有的YAP样本属于单倍群D.总体来 说,YAP+在人群中的比例存在着明显的族 群差异和地理差异. 在日本人和部分藏缅人 群中比例较高; 在华南直至东南亚广泛分布 且在部分人群中有较高比例; 在华北, 东北 以及北亚人群则比例极少甚至没有. 真正的 D*出现在安达曼人,东南亚人群以及藏缅 语人群中, 阿尔泰人中也有少量来源不明的 D*.从单倍群下各个支系来看,D1-M15的 分布区域最宽.它分布在所有东南亚人群, 华南人群, 藏缅语人群中, 一直向东分布到 山东,朝鲜半岛,日本北海道的旭川市;向 北一直分布到辽宁, 内蒙古, 外蒙古的人群 中.D2则仅分布在日韩两个人群中,此外 仅在台湾汉族中观察到1例.D3仅在藏缅语 人群以及临近的人群中观察到.根据此列 表,使用Sufer软件绘制了各支系的分布图 (图1).表1中以下编号人群在地图中被排 除:10,32,113-115,145,161,163.图 1表示YAP在东亚人群的分布.由于唯一已 知出现单倍群E的人群161被排除,因此此 图近似等于单倍群D的分布. 图2表示D以下 的各个亚单倍群在东亚的分布.
表1 Y染色体YAP+在东亚的分布
Tab.1 Distribution of Y chromosome YAP+ in East Asia
表格长度超过排版标准,请从本刊网站下载:
http://COMonCA.org.cn/PDF/2008/COMONCA02-011S1.xls
三,单倍群 D 在东亚各个区域的 人群中的分布及其意义
各个人群中YAP及其支系的分布的详
细数据在表1中,下面将按照地理区域来讨 论其分布. 1.日本人,韩国人(No.1-33) D2- M55几乎局限在日本和朝鲜半岛, 这表明它是D*迁徙到这一地区之后才产生 的.D2- M55的比例在日本南北呈U字型分 布: 在琉球居民和北海道的阿努伊人中比例 最高, 在关西至九州一带最低. 这反映了了 来自大陆的弥生人的移民历史, 这一点翰墨 博士在他的经典文献《日本人的双重起源》 中已经有详细描述[1].D2,D*和D1在朝鲜 人中也有一定比例的分布,从STR结构图 看,朝鲜族的D*并不是与其他人群共享的 单倍型. 我们注意到, 在日本人中没有发现 真正的D*. 由此看来, 朝鲜族中的D在最初 定居这里的人群中就已经存在, 而不是来自 晚近的历史事件.特别要考虑到古老的C3 在历次报道的朝鲜族样本中均占到约10% 的比例,比在日本人中的比例大得多. D1 在北海道旭川市日本人中的出现具 有重要的意义[54].北海道是阿伊努人的世 居地.而在日本其他地区均未发现 D1 的存 在,因此我们可以认为 D1 伴随 D*来到日 韩地区,之后 D*诞生了 D2,而这些 D1 是 日本最早的现代人的遗存. 2.华北,西北少数民族以及中亚人群 (No.34-112) 单倍群D在东北亚大陆(如俄罗斯的埃 文基人)几乎不存在,在中国东北地区也非 常罕见. 但是在蒙古族的各组样本中均有低 频的存在,在0%-6%之间.表1中样本65 来自蒙古国乌兰巴托,同时存在D1和D3. 由于D3主要存在于西藏,这可能反映了历 史时期蒙古与西藏的联系. 甘肃宁夏两省的少数民族也含有一定 比例的D,部分人群中超过10%,而青海互 助县的土族中的D比例更高,总体来说比例 与相对藏区的距离呈反比, 这反映了来自藏 区的人群交流. 肃南至宁夏一带有高比例的 D,与历史上这一代是氐族和党项羌活动的 地区是相符的. 现在这两个古代人群的后裔 均含有一定比例的D.鉴于DYS392=7这个 突变与单倍群D3的特殊联系,文献[3]提供 的 附 件 中 , 表 1 的 人 群 75 有 两 例 样 本 的 DYS392=7属于单倍群D,他们有可能属于 D3.后面将继续详细讨论这一点. 新疆,中亚的人群中也有小比例的 D(总体为2.8%).文献[1]的数据说明,维吾 尔族中有D3,文中也提到,萨彦岭-阿尔泰 地区的阿尔泰人中的D属于D*.文献[26]也 表明阿尔泰人中的某些人群中有高比例的
74
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 D*(非D1,见表1的99-102),但是邻近人群 如哈卡斯人(Khakassians),绍尔人(Shors)却 没有发现.这里的D属于晚近的来自西藏的
动态评论
基因交流还是远古人群的遗存, 有待于利用 STR来进一步详细分析.
图1 Y染色体YAP+在东亚的频率分布 Fig1 Distribution of Y chromosome YAP+ in East Asia This figure is nearly equal to the distribution of haplogroup D in East Asia, because the only population with known Haplotype E in Tab. 1 (No.161) was excluded from this figure. The following data were also excluded: No.10, 32, 113, 114, 115, 145, 161, and 163. Some reports about Chinese ethnic minorities only test the YAP (DYS287) locus. See detail in Tab. 1.
图2 已知的单倍群D1,D2,D3在东亚的分布 Fig.2 Distribution of known Haplogroups D1, D2, and D3 in East Asia 75
FOCUS
COM. on C. A. 2:e11, 2008
3.各地汉族,苗瑶语人群以及侗台语人群 (No.113-271) 总体来说, 汉族中D的比例较少, 在0% -5%之间.在靠近藏区的甘肃,四川则会达 到较高的10%的比例.从文献[65]中D3的 STR网络结构图看,甘肃汉族的绝大部分 D3与藏族的样本共享相同的单倍型,表明 这是来自历史时期人群的融合. 同时, 在部 分地区汉族人群中(比如江苏,山东,详见 表1),也存在少量的D*和 D1.在文献[65] 中D1的STR网络结构图上,汉族的样本有 自己独特的STR单倍型.而这些地区远离 D*高频分布的地区(西藏,东南亚).因此, 各地汉族人群中的D应视为人群的一种遗 传背景存在. 苗瑶语人群以及侗台语人群中 的D*和 D1的大量存在也支持这一点. 单倍群D普遍存在于苗瑶的各个支系 中(0%-10%),在某些支系中D1超过50%, 这应该是迁徙过程中的遗传漂变引起的. 苗 瑶语人群在不断迁徙的过程中, 分化出众多 的支系.在整个苗瑶语人群中比例不大的 Y-SNP单倍群,比如O3a3b-M7,在某些支 系中占到接近100%的比例(黔南州瑶麓的 青裤瑶和茂兰的长衫瑶, 参见文献[87]). 云 南境内的苗瑶语人群也应该经过了这样过 人群的隔离也是引起遗传漂变的原因之 程. 一.我们也可以看到,茶山瑶(拉珈人)居住 的广西金秀县有大瑶山区, 平地瑶居住的 广西富川周围有九嶷山, 萌渚岭, 这些都是 有效隔离人群的地理因素. 畲族的几组数据似乎有些矛盾的地方. 文献[41]检测到YAP+(DYS287)在三个人群 中的比例都很高(近20%).文献[42]的样本 来自同一个县, 检测到的比例为零. 其他的 4组数据也没有检测到YAP+. 台湾原住民各支系以及海南黎族(除了 群体229)中没有发现D,这可能源于起源之 初的人群本身的结构, 长途的迁徙或长期地 理孤立引起的遗传漂变. 但是在大陆上几乎 所有侗台语人群中(包括分化较晚的民族, 如侗族[65],水族[65]等)都出现了D*和D1, 而且在文献[65]中D*和D1的STR网络结构 图上, 侗台语人群的样本有自己独特的STR 单倍型,这说明D也应视为侗台语人群的一 种遗传背景存在. 4.东南亚以及临近的岛屿区域人群 (No.272-305) 石宏等(2008)的论文[65]的重要意义之 一, 就是第一次阐述了安达曼人的父系与亚 洲大陆人群的直接联系.在D*的STR图上, 安达曼人的样本与大陆上的侗台语人群相
76
连. 在现在的侗台语来到东南亚的大陆地区 之前, 这里主要居住着南亚语系的人群. 因 此我们有理由期待看到更多的南亚语人群 (比如柬埔寨人,越南人以及其他孟高棉语 人群)中的D样本以及他们在STR网络结构 图中处于较为接近中心的位置. 但是这一点 没有体现在现有的数据中,文献[65]中也没 有此类数据. 造成这种现状的原因可能有两 个: 1,人群分化历史的差异.侗台语人群 的祖先可能是自东南亚迁往华南的最古老 的人群之一, 因此他们保留了更多的单倍群 D. 云南文山壮族(242), 云南版纳傣族(263) 和泰国清迈泰国人(273,与274一起出现在 文献[65]中)都有较高比例的D*和D1(总和 超过10%). 我们注意到这些人群彼此临近, 都居住在中南半岛中部山区, 地理上的偏远 也有助于人群保留更古老的类型. 而伴随着 水稻栽培引起的人口膨胀,南亚语人群中 Y-SNP主要类型(如O2a-M95)的比例得到增 加,而其他的类型不断被稀释. 2,南亚语人群的样本量过小.历史上 缅甸的南部和东南部, 泰国的南部曾经建立 过很多孟人的国家. 这些地区的样本基本上 还没有分析. 还有长山山脉周围的南亚语人 群也需要大规模采样. 5.中国西南以及藏缅语人群(No.306-400) 中国西南是单倍群D高频存在的区域, 从最初的研究[18]开始,Y-SNP单倍群D与 藏缅语人群的关系就得到了不断的研究. 表 1也包含了国外学者发表的印度东北和喜马 拉雅山以南的藏缅语人群的数据.表1中的 数据表明,单倍群D在整个藏缅语人群的比 例为15.8%. 其他的主要类型还有O3-M122, N*,F*,O2a,C等等,这些单倍群在讨论 整个藏缅语人群的形成过程中都具有重要 的意义,但在本文中我们主要讨论单倍群D 所反映的一些历史事件. 单倍群D在各地区的藏族中的比例较 高,在22.8%-75%之间,平均为45.7%.总 体上讲卫藏附近(拉萨,那曲)的比例较高而 云南藏族的比例较低, 这表明藏区边沿的人 群中D的比例有下降的趋势. 除D2外, 藏族 中 存 在 东 亚地 区 所 有 YAP 的 下 游 支 系 : DE*,D*,D1和D3,这表明他们是东亚最 古老的人群的后裔之一. 与历史上的氐羌人 密切关系的白马藏族和羌族都有较高比例 的D.文献[19]在普米族中检测到了除白马 藏族之外最高比例的D(72.3%, 主要为D3). 但是此外的两组普米族数据中D的比例并
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 不高,平均为33.8%.这表明了人群内部结 构的复杂性. 文献[57]显示尼泊尔以东喜马拉雅山 以南的居民也有较高比例的D,比如锡金人 (Lachungpa),雷布查人(Lepcha).锡金的主 要居民雷布查人原为中国藏族的一部分, 他 们被认为是锡金最早的居民,而锡金人(亦 称布提亚人, Bhutia)是14世纪及以后从西藏 康巴地区迁往锡金的. 德让门巴人历史上也 是藏族的一部分.因此这些人群中D的比例 与藏区接近, 显示了他们历史上与藏族的密 切关系.属于北阿萨姆(塔尼)语支的珞巴族 各部落(阿迪人,阿帕塔尼人,尼西人)中也 有少量的D,根据他们本民族的传说,他们 的祖先是来自西藏的一个小家族. 属于库基 -钦-那加(Kuki-Chin-Naga)语支的南部那加 人(Naga)和属于景颇-孔亚克-波多(JingphoKonyak-Bodo)语支的加若人(Garo)总共超 过800例的样本中,没有检测到D.喜马拉 雅以南, 中缅以西这一大片区域, 居住着大 量的藏缅语人群. 语言学方面的资料尚不完 备, 所以各个语言之间的关系尚不明确. 比 如库基-钦-那加语支下的各种语言,像阿窝 语 (Ao) 与 安 伽 米 - 颇 楚 利 语 (AngamiPochuri),没有明显的证据表明它们属于同 一分支,"之所以把它们放在一起是因为我 们不知道除此之外还可以把它们放在哪 里"[88].从D的角度看,这些人群均含有较 少比例的D, 在整个汉藏语系的人群中,与 汉族的情况最接近. 部分语言学家声称在整 个汉藏语系中汉语与印度东北的藏缅语语 支最接近, 这与遗传学数据有一定程度的 契合. 纳西族中D的比例也较高,与普米族一 样主要为D3,而不是像其他的彝缅语人群 那样基本上为D1.从这一点看,纳西族在 彝缅语人群中的位置特殊, 有待于进一步讨 论.总的来说彝族各支系的D的比例比较大 (部分数据超过15%),而傈僳族,怒族,景 颇族,基诺族和拉祜族中D的比例大都小于 10%,在独龙族,哈尼族中D的比例为零). 这表明, 一些人口较少的分民族由于所处的 地理环境的隔绝, 出现了比较明显的奠基者 效应.结合前述的数据(特别是藏族)可知, 在地理分布上表现为云贵高原的西北部高 (藏族,普米族和纳西族),西南部(景颇族) 和东南部(彝族)略低, 中部更低, 东部(百越 系人群)和南部(南亚语人群)最低.文献[15] 对此作了很详细的分析, 结论认为这是藏缅 语人群南下, 与南方的南亚语人群和侗台语 人群相融合的结果.
77
动态评论
白族和土家族处在藏缅语人群分布的 边沿地区, 可能由于遗传漂变的原因, 他们 中间D的比例比较少. 汉藏语系各语支人群D的分布与语言 学分类之间存在一定程度的对应关系. 除了 共同拥有高比例的O3外,我们可以从以下 遗传学的数据看到藏缅语下各语支的明显 差异.从D的数据看,羌语支的普米族中D 比例极高,藏语支人群中的D普遍高频(接 近50%),彝缅语支人群比例较低(~10%), 而印度东北的那加语支和波多语支人群(除 了最东边的景颇族)中没有D.彝缅语支也 存在一定比例的N*[1,3]和F*[13,64], 则在 其他语支人群中是极少的(其他的部分文献 不检测这些位点).各组汉族数据中也有 10%-15%的N[1,12,13,47,75,76],这代表了 与彝缅语支人群什么样的联系还需要进一 步 分 析 . 此 外 文 献 [64] 表 明 苦 聪 人 拥 有 52.4%的F*,同时考虑O3的话,从现有的 Y-SNP数据看确实与拉祜族拥有最亲密的 关系[13].与彝族直系祖先乌蛮接触过的僰 人也有高比例的F*(10/32,31.3%, [23]). 这表 明彝缅语支人群中存在与其他藏缅语人群 不同的成分. 总结印度方面的线粒体DNA数据以及 以上分析,我们推测单倍群D迁徙路线如下 (图3):D诞生在印度与缅甸之间,然后沿海 岸线向东扩散, 后期进入东南亚和华南内陆 地区, 甚至华北地区. 其中一部分沿海岸线 一直迁徙到现在的日韩地区,之后D*诞生 了D2.另外一部分迁徙到青藏高原边沿, 有可能在末次冰盛期期间也一直生活在那 里.后来其中的D*先后发生了DYS392=7 和D3-P47的突变.在晚近的历史时期,D3 在青藏高原临近区域内部扩张, 可能也伴随 少量D*向中亚和北亚扩散.
图3 Y染色体单倍群D及其分支可能的起源和扩散 途径 Fig.3 Possible origin and diffuse route of Y chromosome Haplogroup D and it's sub-clades
需要注意的是, 由于年代极其古老, 单
FOCUS
COM. on C. A. 2:e11, 2008
倍群D的诞生地可能仍然难以确定,就像超 单倍群DE那样.在整个现代人的迁徙过程 中, 东南亚是重要的中转站, 这里必然发生 过很多次大规模的人口替代. 而在东亚, C, F*, N*以及O这些单倍群可能都有过多次人 口扩张事件, 其中复杂的人群融合过程尚不 清晰, 其间气候也曾发生剧烈变化, 因此早 期迁徙的痕迹难以确定. 单倍群D应视为东亚人群的遗传背景 存在, 并不仅仅是基于它在东亚特殊的分布 状态,也基于与它伴随的mtDNA单倍群. 我们知道,安达曼人的母系全部属于M系 [11],而在东亚大陆(中国),藏区(Y单倍群 鄂伦春族(Y单倍群C D极高频)和鄂温克族, 极高频,[3])生活的内蒙古东部也正是M系 比例最高的地方([89]图4).因此,早期Y单 倍群C和D伴随的母系应是M超单倍群.此 外,M超单倍群下的M9a在日本和藏族人群 中高发,而它的亲缘单倍群E则在东南亚岛 屿地区高频出现, 也支持M系人群作为这一 地区人群的遗传背景的观点[90].
图4 线粒体单倍群M在中国的分布 Fig.4 Distribution of mtDNA haplogroup M in China (from[89])
我们可以看到D3出现在以下群 [65]的数据, 体中: 蒙古的蒙古族, 维吾尔族, 云南普米 族, 西藏, 纳西族, 摩梭人.文献[65]中D3 的STR网络结构图说明:藏族中的D3拥有 绝大部分单倍型并拥有独特的分支, 蒙古族 和维吾尔族的样本从藏族中最主要的单倍 型分出, 汉族的样本绝大部分与藏族分享单 倍型,其他藏缅人群(即纳西族和普米族)的 样本的主要单倍型与藏族共享并拥有一些 独特的细小的分支. 以上的分布图景契合于历史的记录. 后 文将仔细论证,现在拥有D3的人群,均与 一个远古的氏族有关,即藏族古代六氏族: 斯 Se( 塞 / 色 / 赛 ) , 穆 Rmu( 莫 / 木 / 牟 ) , 董 Ldong(顿/桐/洞/党), 冬Stong(东), 札Dbra(惹 /恰/查)和珠Vdru(楚)中的董氏族(据藏族古 代史书《柱间史》,《贤者喜宴》等).经 过学者考证, "董"即是"党项","塞"则是"吐 谷浑(色阿柴)","东"指苏毗,"穆"指象雄 [91,92].不过,石硕教授在《藏族族源与藏 东古文明》中,经过详细的论证认为,"穆" 氏族最初居住在今青藏高原东缘地区的 川西高原一带,"恰"氏族是在今西藏腹心 地区.并且敦煌古藏文写卷中关于"穆", "恰"联姻的记载就隐含在"猕猴与罗刹女 结合衍生藏人"的重要祖源传说[93]. 不过 这可能并不矛盾,"穆"部落在传说时代位 于藏东而在有文献记载的时代出现在阿 里地区.本教文献有提到,"穆族王化身 为大神鸟穹腾空而起, 盘旋良久后降落在 象雄卡佑"[92],这表明穆族可能是外来 的.此外,象雄可能统治过从阿里到康区 之间的所有区域[94].
四,单倍群 D3 的分布
文献[14]中指出, DYS392=7是一个特 殊的突变,文献[14]中的DYS392=7的样本 全部属于D3.而文献[65]显示,DYS392=7 完全属于D*或D3,仅2例例外地属于藏族 D1(可能是由于平行突变的原因). 藏族的28 例D*样本中,有17例的DYS392=7,而D3 中绝大部分(319/328,97.3%)的DYS392=7. 这表明一个事实,D3与藏族中的D*有最密 切 的 亲 缘 关 系 , D3 产 生 于 DYS392=7 的 D*[65]. 根据这一结论和文献[3,13]提供的STR 数据, 我们判断以下人群中的YAP可能属于 D3而肯定属于D:内蒙古的蒙古族, 宁夏回 族, 兰州汉族, 阿坝羌族.结合表1和文献
五,纳西族与藏族在族源和文化 上的亲密关系
赵心愚先生在 《纳西族与藏族历史关系 研究》[95]一文中,对纳西族与藏族的亲缘 关系方面的研究进行了概述(pp15-24), 按照 大意引用如下: P4: ……1922 年 , 美 籍 奥 地 利 学 者 JF洛克(Joseph F.Rock)受美国农业部派遣 来到中国,到丽江后长期居留此地,直到 1949年. 洛克在丽江接触到东巴文化后, 便 大量收集东巴经, 并着手翻译研究……开始 在西方被称为"纳西学研究之父". P5: ……在《中国西南古纳西王国》一 书中, 明确指出藏族人中有一部分与羌人有 关, 同时又指出纳西族原是西藏东北羌人的 一支, 这就强调了纳西族, 藏族存在族源上
78
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 的关系……在《中国西南古纳西王国》中, 洛克多次提到 《格萨尔王传》 中与格萨尔王 打仗的姜三赕王. 在研究了东巴经和藏族传 说后他认为,"三赕"(sa-tham)原在西藏,纳 西南迁时带到云南丽江才被称为三 赕;"姜"(Jang)在藏文中可写作liang,是指么 些.因此,洛克在此书中多次强调,"姜三 赕王"即纳西王,《格萨尔王传》的有关内 容与纳西族,藏族间的战争是对应的. P6: ……在对纳西族东巴教的研究中, 洛克也注意探讨东巴教与藏族本教的关系, 在众多研究者中, 他首先明确指出, 东巴教 的创立者东巴世罗实为"藏人本教的创立者 东巴先饶"(都是藏文ston pa gshen rab 的音 译, [96]). 并认为, 东巴教实际上就是本教, 他甚至断言,"么些人固守着自己的传统巫 教, 这种巫教也曾一度流行于西藏, 但后来 被势力强大的喇嘛教排挤出西藏".他在这 里所说的传统巫教, 即指本教与东巴教. 洛 克提出的这些观点很有价值, 至今仍见其影 响. P9: ……1944年,方国瑜先生发表其著 名长篇论文《么些民族考》,此论文为这一 阶段国内学者的代表作之一……提出了么 些源于 《后汉书》 所记南徙羌人的一支—旄 牛羌的著名观点. 他的这一考证结论, 将早 已提出的纳西族源于羌人说具体化了, 在纳 西族研究中至今仍有很大影响, 仍得到不少 学者的认同. 更具有重要意义的是, 方国瑜 先生在论文中既明确指出么些为南迁羌种, 又明确指出藏族为西迁羌种, 这就强调了两 族族源上存在同源的关系. P11: 俄裔作家顾彼得(Peter Goullart) 在这一阶段的西方纳西族研究者中有一定 影响……顾彼得1955年在英国出版 《被遗忘 的王国》 一书中认为, 纳西族与藏族有某种 渊源关系,这种关系不仅是指语言上存在 "语族亲姻关系",而是因为"纳西族的确是 从西藏下来的".他提出的依据是,"在他们 的用象形文字写成的神圣文学中, 提到马纳 萨洛湖和凯拉斯山, 提到牦牛和居住在高山 草地上的帐篷里"."他们称藏族为大哥,称 白族为弟弟". "他们的祖先稀奇古怪地与印 度神殿里的众神有联系, 他们宣称绝大多数 祖先和英雄是从鸡蛋里魔术般地变出来 的".显然,顾彼得的这些看法是听了不少 纳西族传说及东巴经后才有的, 这说明他对 两个民族的关系有过特别的关注.(这些神 话很明显来自藏族族源的"卵生说", 应是与 明代木氏土司向藏区的扩张和移民有关) P12: 英国爱丁堡大学社会人类学系
79
动态评论
的安东尼杰克逊博士(A.Jackson)……在其 著作的第二章, 第三章的内容中, 他还将纳 西族的信仰体系与相邻的汉族, 藏族的信仰 体系进行比较, 分析了东巴教与本教, 藏传 佛教的关系. 杰克逊认为, 纳西族经书中对 立的二物组合"与喇嘛的二元论很相似,很 多这种二元对应的神灵在藏族和纳西族宗 教中都有相同的名字."这指出了纳西族宗 教与藏族宗教的关系.但他又认为,"丽江 的纳西族没有特殊的单独的宗教信仰, 而选 择适合需要的宗教信仰".从他列举的宗教 来看,皆为藏,汉两族的,这实际上就将东 巴教视为是藏族宗教的一支.他还提出了 "纳西东巴曾经是本教徒","纳西族不可能 有细节上与本教如此相近的平行宗教"的观 点. 他甚至认为, 东巴咒语"是从藏族'真言' 中衍生并因袭下来的", "是纳西对藏人咒语 的音译".事实上,杰克逊已把纳西族宗教 完全视为是藏族宗教的转移. 杰克逊的研究 成果中尽管有不少有价值的观点, 但矛盾的 和错误的看法亦有不少, 将东巴教等同于藏 族宗教就是其中之一. P53:……所列的13个藏缅语民族所存 在的猴祖传说……根据本文前面对纳西族, 藏族猴祖传说所作的分析……表明纳西族 与藏族的猴祖传说存在着惊人的一致性, 这 些一致性可帮助我们在更小的范围通过比 较分析探讨两族猴祖传说的关系, 并进而研 究两个民族在族源上存在的关系. 从上面的论证以及相关的文献[94,97] 可知雍仲本教与纳西东巴教都和原始本教 有一定程度的渊源关系. 《藏族族源与藏东 古文明》中对此也有详细论证([93], pp109 -154).
六,纳西族与远古部落"多弥"
在《历史研究》1985年第1期上,蒙墨 先生发表了重要的 《试论汉代西南民族中的 "夷"与"羌"》 一文, 在"泛羌论"的大背景下, 提出了"夷系民族论". 文中通过对大量文献 的分析, 提出汉代西南民族中除了氐羌濮越 外,还存在另外一个截然不同的民族系统: 即古代被称作"夷"的民族系统, 并认为汉代 西南地区的"夷"系民族应为现在彝缅语支 民族的祖先. 这一观点后来被广泛接受, 比 如1992年出版的 《纳西族史》 中该观点被全 面吸收. 石硕教授在 《藏族族源与藏东古文 明》 称赞"此文对我们从民族系统(族系) 中, 上重新认识青藏高原东缘横断山脉地区的 古代民族具有里程碑的意义"[93,p164] .
FOCUS
COM. on C. A. 2:e11, 2008
关于纳西族《藏族族源与藏东古文明》 一书有以下描述([93],pp165-166):"纳西族 的祖先在 《华阳国志》 中记作"摩沙夷"或 "磨些蛮"….与纳西族有直接传承关系的 "白狼夷"和"牦牛夷"均属于夷系民 族." 因此结论认为纳西族属于夷系民族, 这一点与语言学上的分类一致: 纳西语属于 彝缅语支.但是现有的Y-SNP数据显示,纳 西族中有异于彝缅语人群的成分: 纳西族中 的D绝大部分为D3而彝缅语人群中的D绝 大部分为D1.D1和D3是具有不同人群背景 的单倍群, 因此其中的差异有待下文继续讨 论. 《纳西族与藏族历史关系 赵心愚先生在 研究》[95]一文中,指出纳西族与古代部落 "多弥"的关系.其论证过程按大意引用如 下: P28: 学术意义上的纳西族:纳西族 为新中国建立之后经过民族识别确定的统 一族称,但今云南宁蒗地区的自称"纳日" 的纳西族要求识别为单一的"摩梭族', 今盐 源, 木里一带自称"纳日"的纳西族有不少己 改为蒙古族, 今冕宁雅砻江流域地区的纳木 依人已确定为藏族. 本文下迄20世纪初, 即 1911年辛亥革命,当时这几个地区自称"纳 日","纳木依"的民族都被统称为"么些", 与丽江一带自称"纳西"的称呼一样.因此, 本文仍将几个地区的历史上有着统一称呼 的民族作为一个整体来研究.进入民国后, 尤其是新中国建立后, "么些"这一统称不再 使用. P20: 四川省民族研究所李绍明先生 1981年发表 《康南石板墓族属初探—兼论纳 西族的族源》 一文, 根据考古资料和文献资 料进行分析研究, 认为纳西族源于与牦牛羌 同一族系但不同支的白狼羌, 对似乎已成定 论的纳西族源于牦牛羌的观点发表不同意 见. 这一新看法的提出, 推动了对纳西族族 源问题的进一步探讨. 他还认为, 白狼羌留 在康南地区者后来逐渐融入吐蕃.②实际 上,他已指出两族存在某种渊源关系. P31:在丽江一带的纳西族中,直到现 在仍有这样一种口头传说:他们的祖先原并 不居住在云南丽江, 而是住在北方一个名叫 "多弥"的地方, 后来才沿金沙江等河谷往南 迁徙到达现居住地. 因此, 有人认为纳西族 族源传说中的多弥可能与唐代西北的多弥 部落有关.也有学者依此认为,"多弥"一词 应早己存在. P33:按纳西族丧葬习俗,人死后,要 请东巴(或达巴)将死者灵魂从家中一站一
80
站送往祖先最初的居住地, 送魂路线纳西语 称作"哦般日", 意为灵魂迁徙之路……他们 的送魂路线基本上就是反过来的起祖路 线……四川盐源县左所区纳西族人, 永宁纳 西族,盐源纳西族(自称纳日人),冕宁木里 纳木依人等各地纳木依人接祖先的路线均 指向北方或最终到达昆仑山. 而根据任乃强: 《释巂昆明》[载《羌族源流探索》,重庆 出版社1984年版]一文,昆仑部落即隋唐时 期之多弥羌,多弥是吐蕃征服他们后的称 呼. P36: (关于多弥部落自称与纳西族制自 称的关系的重要论证) 据《新唐书西域传》记载:"多弥,亦 西羌族,役属吐蕃,号难磨.滨犁牛河,土 多黄金."从这一记载中,可知多弥为古羌 人之一部, 当时己臣服于吐蕃. 犁牛河为通 天河异称,近河源,其居住地"滨"犁牛河, 即在今长江上游流经青海玉树州一带. 值得 注意的是这一记载中提到的"难磨"一词. 关 于"难磨",笔者在本节第二部分还要讨论, 这里着重讨论"难磨"与纳木依人, 纳日人的 关系.根据以上记载,"难磨"应为多弥羌自 号.任乃强先生考证"路巴"之"洛"时指出, "路"为"那"的音变,本意为"黑",后借指南 方. "难磨"之"难",古音可读"摊",因而 极可能也为"纳"的音变,本意也为"黑".当 然, 这仅是一种推测. 从语言调查来看, "难" 实际上就是"纳"的特殊读音, 至少在纳木依 人和纳日人中是如此. 有学者在调查材料中 称,永宁纳西族(纳日人)称他们的歌为"纳 刮",意即纳日人的歌.但在中国科学院民 族研究所的调查材料中, 称永宁纳日人的歌 当地人叫做"南刮".这说明,纳日人对"纳" 的发音与"南"基本相同. 不仅纳日人发音是 如此, 纳木依人的发音也是如此, 其自称发 言听起来即为"南木依". "难"与"南"音同, 可以认为"难磨"实为"纳磨"."难磨"之"磨" 与西夏语中"MO"的记音同,意即"人",与 羌语"人"的读音"MI''也相近.四川木里, 盐源的纳日人称始祖为"纳木',音与"难磨" 相同,其意为"黑人".在纳木依语中,"纳 木依"一词后两字快读可读为"纳米",如重 音放在"纳"上, 也可读为"纳木". 在含义上, 纳木依一词亦指" 黑人". P38 :除了这些明确记载外,汉文史 籍中还有一些记载也可能是有关多弥的. 《隋书》在记附国时有这样一段记载:"附 国……东北连山,绵亘数千里,接于党项. 往往有羌:大小左封,昔卫,葛延,白狗, 向人,望族,林台,春桑,利豆,迷桑,婶
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 药, 大砍, 白兰, 叱利摸徒, 那鄂, 当迷…… 并在深山穷谷, 无大君长. 其风俗略同党项; 或役属于吐谷浑,或附附国."《北史附国 传》 亦有基本相同的记载. 中, 有学者认为, 《隋书》所记的"当迷"即唐代的多弥.从 记音和地理位置来看, 当迷应是多弥的 异译.《旧唐书吐蕃传》中有"当弥道", 此"当弥"王忠先生考证后认为即多弥.又, 《新唐书吐蕃传》记载:"吐蕃令曩骨委书 塞下,言:'论莽热,论泣热皆万人将,以赞 普命,谢都督刺史:二国有舅甥好,昨弥不 弄羌,党项交构二国,故失欢,此不听,唐 亦不应听."陈宗祥先生认为,这一记载中 昨弥即多弥的 的"不弄"为白兰的一种译法, 异译.从记音来看,"不弄"可能为"白兰" 或"白狼"的异译, 但从多弥早己役属吐蕃来 看,此昨弥当另有所指,不是指多弥. ……多弥号难磨, 而难磨之"难"(Nam) 正是吐蕃时代藏文文献中提到的"南国"之 "南"(Nam)的对音. 因此一些中外学者认为, " 南 " 国 即 藏 文 文 献 记 载 中 的 多 弥.……"磨"(mo)在西夏语中为"人"的意 思,多弥与党项同处一地,同为羌人,在多 弥语中"磨"也当为"人"的意思……实际上, "南国"之"南"与"磨难"之"难"都是"纳"的一 种读音,其意本为"黑"."难磨"一词为一完 整概念,即指"黑人".这里的"黑人"不是指 肤色, 服饰, 而是一个族系概念. 可以认为, 多弥人的"号"也就是自称的含义为"黑人". P40: 据藏族著名学者巴卧祖拉陈哇 所撰《贤者喜宴》记载,松赞干布时代吐蕃 周围有"四方面王"(或称"四外邻邦"),即囊 巴(Nam-pa)财库王,泥婆罗(今尼泊尔)铜 王,苏毗铁王及门地(今西藏门巴)娱乐 王……, 他们收集赋税上献, 因此他们隶属 于属民之中."这一记载中的"囊巴"当为难 磨的另一种译法,"囊"与"难"对音,"巴"与 "磨"则均指人.所以,囊巴财库王当为被吐 蕃征服后的多弥部落首领……法国著名藏 学家RA石泰安(Stien)在研究中注意到, 与 吐蕃王朝赞普通婚, 结盟的家族集团中有一 大家族称那囊氏(SNa-nam),他指出这一家 族归顺后与许布,蔡邦,贝农"变成了吐蕃 四大贵族家族. "…… 这是融入吐蕃的多弥 人的线索. 由以上论证可知,摩梭人,纳日人,纳 木依人和纳西族均是古代部落多弥(自称难 磨)的后裔.
动态评论
族和穆族
《隋书党项传》载:"党项羌者,三苗 之后也.其外有宕昌,白狼,皆自称称猴 种."[98] 在《北史党项传》中,也有相同 的记载.陈宗祥先生认为,白狼即《后汉 书西羌传》记载的青海黄河南岸羌人"卑 湳"部落之一部[99].白狼与纳西族族源的 关系问题不少学者己有专门论述. 方国瑜先 生在《么些民族考》中,早己根据史籍记载 指出白狼为牦牛羌的一支, 摩沙夷亦为牦牛 羌的一支,二者同种[100].李绍明先生在 一篇论文中认为 "白狼羌一部分后来沿着 金沙江, 雅砻江进入今川滇边境一带, 其后 裔即晋代的摩沙夷以及唐代的磨些以及现 今的纳西"[101] .史籍记载与大量传说等 材料说明白狼人与纳西族确存在族源联系, 为其族源之一, 学术界对此的看法也基本一 致. 对于南语,多弥,白兰与董族的关系, 《多弥史钩沉》[102]中提到:在敦煌发现 的古藏文卷子中,有一份是写于公元8-9世 纪的, 用古藏文书写的一种近似藏语而尚未 确知的语言,经托玛斯研究,定名为 "南"(Nam)语. 为此, 他撰写了一部长达400 多页的专著: 《南,一种汉藏边境的古代语 言》".这个南木语抄写本经藏学家陈宗祥 教授多年深入研究, 译为汉文, 整理成书, 命名为 《南语—汉藏民族走廊的一种古代羌 语》出版."事实上,白兰应为南部的一支, 即 sphrom( 意 为 " 白 色 " , 音 译 为 " 不 弄")Nam(Lam,南)部族. 即是说, 多弥(南国) 与之邻近的白兰均源于汉代以来西羌中的 卑湳等羌…… 为什么汉文史籍称"南"部族 为"多弥"呢 笔者认为, 据敦煌发现的吐蕃 历史文书纪年中, 有公元702年"冬, 赞普驻 于墀寨,于'南木东兆木'(Nam LDong prom) 地方……主持多思麻之冬季集会议盟会." 而所谓的"LDong prom", 南语意为"白董族" 之 意 . ……正 如 法 国 著名 汉 学 家 石泰 安 (RAStein)所说, 南族也往往与董族联接在 一起称Nam-LDong. 因此, "多弥"似可还原 为"LDong mi"即董弥;mi为一般藏缅语族 称之为人之意,也就是"董族人"之意" 陈宗祥教授在上世纪80年代初发表的 [103]提到,""不弄"即"白兰",其藏文 对应词就是《格萨尔王传》中 "格萨尔" 总有一个的前缀Phrom." 在新近发表的 一篇文章中[104] 陈宗祥教授又论证了《隋 书附国传》中的婢药就是附国.附国的首 府在邓柯县. 邓柯县也叫邓玛县, 现在分属
七,藏族"原始四氏族"中的董
81
FOCUS
COM. on C. A. 2:e11, 2008
德格县, 石渠县. 今德格县南有白玉县(dbat yul),即"附国南有薄缘夷"中的薄缘[103]. 而我们知道, 德格县被认为就是董族后裔的 岭格萨尔王的诞生地. 文献[103] 中也提到,"《南语写卷》1 号146行说,"南家与邓玛(vtong ramg)一起 掌管部落"." 文献[105]提到:史诗《格萨 尔王》中有勇猛仅次于格萨尔王的邓玛大 将. 在有关格萨尔王生世的叙述中, 经常提 到格萨尔王出生在康区"邓", "岭"两部落的 "岭"部落.可见, "邓"或者"邓玛"先是一 部族或一地区的名字(邓域),后来变为人 名.经论证,认为邓玛大将是邓萨霍王的 后裔.而[106]中三国至西晋的详图中,四 川西北部出现的"邓麻, 马儿敢"中邓麻的地 域与邓玛重合, 而读音完全相同, 当即邓玛. 而据[95]p26论述:据《新唐书西域传》记 载:"多弥,亦西羌族,役属吐蕃,号难磨. 滨犁牛河,土多黄金."从这一记载中,可 知多弥为古羌人之一部,当时己臣服于吐 蕃.犁牛河为通天河异称,近河源,其居住 地"滨"犁牛河, 即在今长江上游流经青海玉 树州一带.而格萨尔王与邓玛大将出生的 "邓", "岭"两部落的居地就在通天河雅砻江 上游之间的德格县,石渠县一带 [102,103,105].多弥与邓玛的居住地也是重 叠的. 据此, 我们认为多弥和邓玛可能是同 一个部落名的不同音译. 文献[107]提到:格萨尔史诗,藏语中
称为"格萨尔的故事",简称"仲"或"岭仲", 意译即"故事","岭国的故事"…. 现已出版 的《世界形成》,《董氏预言授记》,《天 界篇》等书,讲述了格萨尔祖先"董"氏族的 来源,世系和传承; 《诞生篇》和《赛马篇》 等书,讲述格萨尔诞生和"董"的"父系三兄 弟"从卫藏辗转迁徙到黄河源头,并占领其 地的经历…. 这个史诗唱段告诉我们,"董" 氏族的发展线索是:古代六氏族——"穆布 董"——岭六部——三十个众兄弟.我们从 藏文历史文献中终于找到了"岭巴"这氏族 (部落).它原来是穆布董氏族中的"尊者六 系"之一,并不像格萨尔史诗中描写的那么 强大.但"岭巴"这个氏族(部落)在历史上的 确存在,它的确有"古代六氏族"——"穆布 董"——"岭巴"的源远流长的历史发展线 索. 在现在的德格县拥有与格萨尔王相关 的最多的遗址和传说. 很多学者认为, 格萨 尔王真有其人, 真有其事. 在康区特别是德 格地区, 人们都深信岭格萨尔王确有其人, 他就是原邓柯林葱家族的祖先, 其出生地在 今德格(原邓柯)县阿须乡雄坝吉苏雅格康 多.产生于康区特别是德格地区的手抄本, 木刻本, 以及民间说唱形式的艺人最多, 最 权威.这里有全藏区最权威版本的"林葱木 今德格县, 石渠县境内有大量与 《格 刻本". 萨尔王传》中关联的遗迹[108-111].
图5 现代族群与西藏古代董氏族的 关系
Fig.5 Relations between contemporary populations with ancient tribe "Ldong"in Tibet
82
兰海:Y 染色体单倍群 D 在东亚的分布及其意义
动态评论
图6 青藏高原历史上的各个政权/部族 修改自[106]唐详图(669年) Fig.6 Ancient regimes and tribes in Tibet Modified from [106] Detail map of Tang Dynasty(669A.D.)
根据以上论述,远古至今的各部落/人 群的演化关系如图 5 所示.需要说明的是, 在历史时期,各部落不断分化,迁徙,与当 地人群的融合是不可避免的, 因此图中的箭 头仅表示一定程度的继承关系. 我们可以看 到,藏族,纳西族和普米族这些拥有 D3 的 人群, 均与一个远古的董氏族有关. 不过可 以肯定的是, 并非所有的 D3 均来自董氏族. D3 的年代在文献[14]中超过 1 万年.由于 真正的 D*是较少的,因此我们可以谨慎地 推测, 白马藏族中(50%为非 D1 的 D*)也有 较大比例的 D3.而历史上没有其他地区藏 族大规模融入白马藏族的记录. 文献的记录 显示, 现在的藏族正是卫藏地区, 阿里地区, 康藏地区, 安多地区以及青海地区等历史上 多个部落群体融合而成的. 为了清楚表示图 5 中的演化关系,在 [106]唐详图(669 年)的基础上进行修改,加 入图 5 中不同历史时期的各个部落/政权, 结果如图 6 所示. 大致的地域范围参考了才 让教授的《吐蕃史稿》[112]. 居住在喜马拉雅以南门隅, 洛隅地区的 门巴族和珞巴族与藏族在族源上有亲密的 关系.据文献[113]: P22:在珞巴族诸部落中,有诸多反映 珞巴族和藏族的祖先是亲兄弟的传说," 太 阳母亲和月亮父亲生下了阿波崩戎和阿波
83
达尼. 阿波崩戎是哥哥, 他的子孙后代繁衍 发展成了藏族. 阿波达尼是弟弟, 他的子孙 后代繁衍发展成了珞巴族.门巴族人常 说:"我们同藏族,珞巴族是同宗同祖吃一条 江水的兄弟". 共同拥有的猴子变人的传说, 深刻地昭示了西藏高原上珞巴族, 门巴族和 藏族紧密的文化联系和对同一血缘祖先的 认同心理." P23:《门隅教史》叙述: ……其后, 称猴之后裔世代演化,成斯,穆,董,冬四 大族系…. 穆分化成的归,列,查等八个姓 氏为:俄布,修布,雄,门,嘎,噶,虐, 恩兰等.其中的"门",指的是门巴先民. 从表 1 我们可以看到(No.331-338),门 巴族中有较高比例的 D,而同为"穆氏族" 后裔的珞巴族则含有极高比例的 O3.结合 O3 在其他汉藏语人群都有较高的比例以及 其他单倍群(如 N*)的大量存在的事实,我 们可以确定, 自远古时代以来, 青藏高原上 的氐, 羌, 夷以及原始六氏族等部落都已经 是深度融合的人群.
参考文献
1. Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S(2006) Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes. J Hum Genet 51:47-58. 2. Wen B (2004) Y chromosome, mtDNA variation and the genetic structure of East Asians. PhD Thesis of Genetics,
FOCUS
FudanUniversity. 3. Xue YL, Zerjal T, Bao WD, Zhu SL, Shu QF, Xu JJ, Du RF, FuSB, Li P, Hurles ME, Yang HM, Tyler-Smith C (2006) Male Demography in East Asia: A North-South Contrast in HumanPopulation Expansion Times. Genetics 172:2431-2439. 4. Wells RS, Yuldasheva N, Ruzibakiev R, Underhill PA, Evseeva I, Blue-Smith J, Jin L, Su B, Pitchappan R, Shanmugalakshmi S,Balakrishnan K, Read M, Pearson NM, Zerjal T, Webster MT,Zholoshvili I, Jamarjashvili E, Gambarov S, Nikbin B, DostievA, Aknazarov O, Zalloua P, Tsoy I, Kitaev M, Mirrakhimov M,Chariev A, Bodmer WF (2001) The Eurasian heartland: a continental perspective on Y-chromosome diversity. Proc Natl Acad Sci USA 98:10244-10249. 5. Karafet T, Xu L, Du R, Wang W, Feng S, Wells RS, Redd AJ, Zegura SL, Hammer MF ( 2001) Paternal population history of East Asia: ources, patterns, and microevolutionary processes. Am J Hum Genet 69: 615-628. 6. Kivisild T, Rootsi S, Metspalu M, Mastana S, Kaldma K, Parik J, Metspalu E, Adojaan M, Tolk HV, Stepanov V, Glge M, Usanga E, Papiha SS, Cinniolu C, King R, Cavalli-Sforza L, Underhill PA, Villems R (2003)The genetic heritage of the earliest settlers persists both in Indian tribal and caste populations. Am J Hum Genet 72:313-332. 7. Qian Y, Qian B, Su B, Yu J, Ke Y, Chu Z, Shi L, Lu D, Chu J, Jin L (2000)Multiple origins of Tibetan Y chromosomes. Hum Genet 106:453-454. 8. 文波,石宏,任玲,奚慧峰,李开源,张文逸,宿兵,司世恒,金力, 肖春杰(2003) Y染色体,线粒体DNA多态性与云南宁蒗摩 梭人的族源研究.中国科学(C辑)33:375-384. 9. Su B, Xiao J, Underhill P, Deka R, Zhang W, Akey J, Huang W,Shen D, Lu D, Luo J, Chu J, Tan J, Shen P, Davis R,Cavalli-Sforza L, Chakraborty R, Xiong M, Du R, Oefner P,Chen Z, Jin L (1999)Y-chromosome evidence for a northward migration of modern humans into Eastern Asia during the Last Ice Age. Am J Hum Genet 65: 1718-1724.9 10. Du H, Zhang L, Zhou B, Zhang HJ, Liang WB, Shen YH (2006) A Minisequencing Technique for Genotyping 12 Y-SNP and it's Population Data for Chinese Population in Sichuan. J Forensic Med(Chin) 22:125-129. 11. Thangaraj K, Singh L, Reddy AG, Rao VR, Sehgal SC, Underhill PA, Pierson M, Frame IG, Hagelberg E (2003) Genetic Affinities of the Andaman Islanders, a Vanishing Human Population. Curr Biol13: 86-93. 12. Kayser M, Brauer S, Weiss G, Schiefenhvel W, Underhill P, Shen P, Oefner P, Tommaseo-Ponzetta M, Stoneking M (2003)Reduced Y-chromosome, but not mitochondrial DNA, diversity in human populations from west New Guinea. Am J Hum Genet 72: 281-302. 13. Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, EdmondsCA, Chow CE, Lin AA, Mitra M, Sil SK, Ramesh A, Usha Rani MV, Thakur CM, Cavalli-Sforza LL, Majumder PP, Underhill PA (2006) Polarity and temporality of high-resolution Y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists. Am J Hum Genet 78:202-221. 14. Gayden T, Cadenas AM, Regueiro M, Singh NB, Zhivotovsky LA, Underhill PA, Cavalli-Sforza LL, Herrera RJ (2007)The Himalayas as a directional barrier to gene flow. Am J Hum Genet 80:884-894. 15. Wen B, Xie X, Gao S, Li H, Shi H, Song X, Qian T, Xiao C, Jin J, Su B, Lu D, Chakraborty R, Jin L (2004)Analyses of genetic structure of Tibeto-Burman populations reveals sex-biased admixture in southern Tibeto-Burmans. Am J Hum Genet 74:856-865. 16. Su B, Xiao C, Deka R, Seielstad MT, Kangwanpong D, Xiao J, Lu D, Underhill P, Cavalli-Sforza L, Chakraborty R, Jin L(2000) Y chromosome haplotypes reveal prehistorical migrations to the Himalayas. Hum Genet 107:582-590. 17. Chen J, Li H, Qin ZD, Liu WH, Lin WX, Yin RX, Jin L, Pan SL (2006)Y-chromosome Genotyping and Genetic Structure of Zhuang Populations. Acta Genet Sin33:1060-1072. 18. 钱 亚 屏 , 初 正 韬 , 卫 灿 东 (1999) 中 国 云 南 省 五 个 民 族 DYS287 位 点 多 态 性 的 调 查 . 中 华 医 学 遗 传 学 杂 志
COM. on C. A. 2:e11, 2008
16:381-382. 19. 石宏,董永利,李卫翔,杨洁,李开源,昝瑞光,肖春杰(2002)中 国云南25个少数民族Y染色体DYS287位点的地理多态性. 中国科学C辑(生命科学) 32: 373-378. 20. 董永利,高路,杨晓霞,文波,杨智丽,唐文如,程宝文,郑冰蓉, 李开源,肖春杰(2005)白族群体间的Y染色体和线粒体DNA 多态性的分析. 遗传学报 32:450-456. 21. 李冬娜,应大君,区采莹,陈路,符生苗,谢协驹,陈峰(2003)中 国海南岛三个黎族支系DYS287,DYS19的多态性研究.遗 传 25: 5-8. 22. 李永念,左丽,文波,柯越海,黄薇,金力(2002) 中国布依族 人的起源及迁移初探-来自Y染色体和线粒体的线索.遗传 学报29: 196-200. 23. 于敏,张咏莉,陈峰,薛雅丽,于旸,马琳琳,黄小义,刘岸,史榕 茜,吕芙蕖,黄承滨,张贵寅, 李璞,傅松滨(2002)中国6个人群 中Y染色体15个双等位基因标记变异频率分布及单体群分 析.遗传学报29: 283-289. 24. 石宏,李易,罗虹,吉学平,宿兵(2006)云南僰人源流的父系 和母系遗传研究. 动物学研究27:449-455. 25. 董永利,杨智丽,石宏,高路,鲁靖,程宝文,李开源,昝瑞光,肖 春杰(2004) 云南18个民族Y染色体双等位基因单倍型频率 的主成分分析. 遗传学报 31:1030-1036. 26. Khar'kov VN, Stepanov VA, Medvedeva OF, Spiridonova MG, Voevoda MI, Tadinova VN, Puzyrev VP(2007) Gene pool differences between northern and southern Altaians inferred from the data on Y-chromosomal haplogroups. Genetika. 43:675-687. 27. Kim W, Shin DJ, Harihara S, Kim YJ (2000) Y chromosomal DNA variation in East Asian population and its potential for inferring the peopling of Korea. J Hum Genet 45:76 -83. 28. 张咏莉,于敏,薛雅丽,陈峰,黄承滨,张贵寅,李璞,傅松滨 (2002)中国福建汉族14个YSNPS位点的研究.人类学学报 21:245-249. 29. Derenko M, Malyarchuk B, Denisova G, Wozniak M, Dambueva I, Dorzhu C, Luzina F, Micicka-liwka D, ZakharovI (2006) Contrasting patterns of Y-chromosome variation in South Siberian populations from Baikal and Altai-Sayan regions. Hum Genet 118:591-604. 30. Lell JT, Sukernik RI, Starikovskaya YB, Su B, Jin L, Schurr TG, Underhill PA, Wallace DC (2002) The Dual Origin and Siberian Affinities of Native American Y Chromosomes. Am J Hum Genet 70: 192-206. 31. 李辉,李昕,杨宁宁,文波,侯井榕,黄兴球,金建中,金力,卢大 儒(2003)遗传和体质分析草苗的起源.复旦学报(自然科学 版) 42: 621-629. 32. 何燕,文波,单可人,黄薇,谢渊,吴昌学,熊晓燕,修瑾,张静, 张小蕾,任锡麟,金力(2003)贵州三都水族Y染色体单倍型频 率分析.遗传25: 249-252. 33. 李辉,杨宁宁,黄兴球,陈莉,金建中,金力,卢大儒(2002)广西 融水北高村人的遗传学和体质人类学特征.中央民族大学 学报 11:108-116. 34. 李辉(2007)分子人类学所见历史上闽越族群的消失.广西 民族大学学报(哲学社会科学版) 29(2):42-47. 35. 李辉,潘悟云,文波,杨宁宁,金建中,金力,卢大儒(2003)客家 人起源的遗传学探索.遗传学报 30:7-14. 36. 谢选华(2004)从Y染色体遗传结构看藏缅语族人群的起 源. 硕士毕业论文,复旦大学. 37. 文普帅,温有锋,席焕久,任甫,高静(2007)西藏藏族DYS287 位点多态性研究.武警医学 12: 89-90. 38. 陈阳,褚嘉佑,俞建昆,于亮,孙浩,林克勤,陶玉芬,史磊,黄小 琴 , 石 铁 流 , 傅 松 滨 (2006) 中 国 28 个 民 族 群 体 Y 染 色 体 DYS287 位 点 的 遗 传 多 态 性 . 中 国 医 学 科 学 院 学 报 28:196-201. 39. Wen B, Li H, Lu D, Song X, Zhang F, He Y, Li F, Gao Y, Mao X, Zhang L, Qian J, Tan J, Jin J, Huang W, Deka R, Su B, Chakraborty R, Jin L (2004)Genetic evidence supports demic diffusion of Han culture. Nature 431:302-305. 40. 张勇,张贵寅,孙艳阳,薛雅丽,杨焕杰,陈白滨,李璞(1998)中 国东北汉族及3个少数民族DYS19和DYS287多态性研究. 人类学报17:237-241. 41. 范启兰,张文峰,郭俊明,叶金花,赖日勇,许春鹃(2002)中国 畲族人群DYS287位点多态性的研究.赣南医学院学报 22: 323-325. 42. 闫春兰,詹金彪,邵靖宇(2004)浙江畲族DYS287DYS440 位点的多态性研究.中华医学遗传学杂志 21: 410-411. 43. 周瑞霞,安黎哲,伊琳,杨亚军,王勋陵,张伟华,郭茜,谢小冬
84
兰海:Y 染色体单倍群 D 在东亚的分布及其意义
(2006)中国甘肃三个特有少数民族回族和汉族DYS287 DYS19的遗传多态性研究.兰州大学学报 (自然科学版) 42(2):38-42. 44. 何燕,单可人,吴昌学,谢渊,李毅,官志忠,任锡麟(2006)贵州 省三都水族DYS287和DYS388基因座多态性分析.贵阳医 学院学报31:400-405. 45. Jin HJ, Kwak KD, Hammer MF, Nakahori Y, Shinka T, Lee JW, Jin F, Jia X, Tyler-Smith C, Kim W (2003) Y-chromosomal DNA haplogroups and their implications for the dual origins of the Koreans. Hum Genet 114: 27-35. 46. Gan RJ, Pan SL, Mustavich LF, Qin ZD, Cai XY, Qian J, Liu CW, Peng JH, Li SL, Xu JS, Jin L, Li H; Genographic Consortium (2008) Pinghua Population as an Exception of Han Chinese's Coherent Genetic Structure. J Hum Genet 53:303-313. 47. 马明义,杨元,肖翠英,张思仲(2007) 四川汉族人群Y染色 体单倍组及其特点的研究.中华医学遗传学杂 志.24:261-265. 48. 董永利,石宏,李卫翔,杨洁,曾卫民,李开源,肖春杰(2002)怒 江大峡谷及下游地区7个云南少数民族YAP位点的多态性 研究.人类学学报 21:249-254. 49. Tajima A, Hayami M, Tokunaga K, Juji T, Matsuo M, Marzuki S, Omoto K, Horai S(2004) Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages.J Hum Genet. 49(4):187-193 50. Tajima A, Pan IH, Fucharoen G, Fucharoen S, Matsuo M, Tokunaga K, Juji T, Hayami M, Omoto K, Horai S (2002) Three major lineages of Asian Y chromosomes: implications for the peopling of east and southeast Asia. Hum Genet 110: 80-88. 51. 俞建昆,孙浩,史磊,钱亚屏,史荔,黄小琴,褚嘉佑(2004)中国 3个民族群体Y染色体上17个双等位基因位点的遗传分析. 中国医学科学院学报 26:537-542. 52. 俞建昆,孙浩,史磊,史荔,钱亚屏,黄小琴,车艳春,褚嘉佑 (2006) 5个中国人群Y染色体上17个双等位基因位点的多 态性分析. 应用与环境生物学报12: 247-250. 53. 黄小义(2004)5个中国人群中Y染色体非重组区双等位基 因多态性的分布.硕士论文,哈尔滨医科大学. 54. Nonaka I, Minaguchi K, Takezaki N (2007) Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms. Ann Hum Genet 71(Pt 4):480-495. 55. Su B, Jin L, Underhill P, Martinson J, Saha N, McGarvey ST, Shriver MD, Chu J, Oefner P, Chakraborty R, Deka R (2000)Polynesian origins: Insights from the Y chromosome. Proc Natl Acad Sci U S A 97:8225-8228. 56. Bender K, Stradmann-Bellinghausen B, Rittner C, Schneider PM (2003) Comparative analysis of short tandem repeats and single nucleotide polymerphisms on the Y-chromosome in Germans, Chinese and Thais. Leg Med (Tokyo) 5:164-168. 57. Chandrasekar A, Saheb SY, Gangopadyaya P, Gangopadyaya S, Mukherjee A, Basu D, Lakshmi GR, Sahani AK, Das B, Battacharya S, Kumar S, Xaviour D, Sun D, Rao VR (2007) YAP insertion signature in South Asia. Ann Hum Biol 34:582-586. 58. Hong SB, Jin HJ, Kwak KD, Kim W (2006)Y-chromosome haplogroup O3-M122 variation in east Asia and its implications for the peopling of Korea. Korean J Genet 28: 1-8. 59. Katoh T, Munkhbat B, Tounai K, Mano S, Ando H, Oyungerel G, Chae GT, Han H, Jia GJ, Tokunaga K, Munkhtuvshin N, Tamiya G, Inoko H (2005) Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis.Gene 346:63-70. 60. Nasidze I, Quinque D, Dupanloup I, Cordaux R, Kokshunova L, Stoneking M(2005) Genetic evidence for the Mongolian ancestry of Kalmyks. Am J Phys Anthropol 128:846-854. 61. Tan S, Yang M, Yu H, Dong Y, Shou W, Zou J, Tang W, Guo Y, Xiao C (2007) Y-chromosome polymorphisms define the origin of the Mang, an isolated population in China. Ann Hum Biol 34:573-581. 62. Zhou R, Yang D, Zhang H, Yu W, An L, Wang X, Li H, Xu J, Xie X (2008) Origin and evolution of two Yugur sub-clans in Northwest China: a case study in paternal genetic landscape. Ann Hum Biol 35:198-211. 63. Li H, Wen B, Chen SJ, Su B, Pramoonjago P, Liu Y, Pan S,
动态评论
Qin Z, Liu W, Cheng X, Yang N, Li X, Tran D, Lu D, Hsu MT, Deka R, Marzuki S, Tan CC, Jin L(2008) Paternal genetic affinity between western Austronesians and Daic populations. BMC Evol Biol 5: 146. 64. Bittles AH, Black ML, Wang W(20007) Physical anthropology and ethnicity in Asia: the transition from anthropometry to genome-based studies. J Physiol Anthropol 26:77-82. 65. Shi H, Zhong H, Peng Y, Dong YL, Qi XB, Zhang F, Liu LF, Tan SJ, Ma RL, Xiao CJ, Wells S, Jin L, Su B(2008)Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biol 6:45. 66. Bhaswar Maity, T Sitalaximi, R Trivedi and VK Kashyap(2004) Tracking the genetic imprints of lost Jewish tribes among the gene pool of Kuki-Chin-Mizo population of India. Genome Biol 6:P1. 67. Cordaux R, Weiss G, Saha N, Stoneking M (2004) The northeast Indian passageway: a barrier or corridor for human migrations Mol Biol Evol 21:1525-1533. 68. Kumar V, Reddy AN, Babu JP, Rao TN, Langstieh BT, Thangaraj K, Reddy AG, Singh L, Reddy BM (2007) Y-chromosome evidence suggests a common paternal heritage of Austro-Asiatic populations. BMC Evol Biol 7:47. 69. 李永念,左丽,文波,黄微,金力(2003)Y染色体单倍型在贵 州苗族人群中的多态性分布.贵阳医学院学报 28:98-105. 70.Underhill PA, Shen P, Lin AA, Jin L, Passarino G, Yang WH, Kauffman E, Bonné-Tamir B, Bertranpetit J, Francalacci P, Ibrahim M, Jenkins T, Kidd JR, Mehdi SQ, Seielstad MT, Wells RS, Piazza A, Davis RW, Feldman MW, Cavalli-Sforza LL, Oefner PJ(2000) Y chromosome sequence variation and the history of human populations.Nat Genet 26:358-361. 71. Hammer MF (1994) A recent insertion of an Alu element on the Y chromosome is a useful marker for human population studies. Mol Biol Evol 11:749-761. 72. Santos FR, Bianchi NO, Pena SD (1996) Worldwide distribution of human Y-chromosome haplotypes. Genome Res 6:601-611. 73. Hammer MF, Spurdle AB, Karafet T, Bonner MR, Wood ET, Novelletto A, Malaspina P, Mitchell RJ, Horai S, Jenkins T, Zegura SL (1997) The geographic distribution of human Y chromosome variation. Genetics 145:787-805. 74. Hammer MF, Zegura SL(1996)The role of the Y chromosome in human evolutionary studies. Evol Anthropol Issues News Rev 5:116-134. 75. Hammer MF, Karafet TM, Redd AJ, Jarjanazi H, Santachiara-Benerecetti S, Soodyall H, Zegura SL (2001) Hierarchical patterns of global human Y-chromosome diversity. Mol Biol Evol 18:1189-1203. 76. Spurdle AB, Hammer MF, Jenkins T (1994) The Y Alu polymorphism in southern African populations and its relationship to other Y-specific polymorphisms. Am J Hum Genet 54: 314-330. 77. Deng W, Shi B, He X, Zhang Z, Xu J, Li B, Yang J, Ling L, Dai C, Qiang B, Shen Y, Chen R (2004) Evolution and migration history of the Chinese population inferred from Chinese Y-chromosome evidence. J Hum Genet 49:339-348. 78. Hammer MF, Horai S (1995)Y chromosomal DNA variation and the peopling of Japan. Am J Hum Genet 56: 951-962. 79. Altheide TK, Hammer MF (1997)Evidence for a possible Asian origin of YAP+ Y chromosomes. Am J Hum Genet 61:462-466. 80. Hammer MF, Karafet T, Rasanayagam A, Wood ET, Altheide TK, Jenkins T, Griffiths RC, Templeton AR, Zegura SL (1998) Out of Africa and back again: nested cladistic analysis of human Y chromosome variation. Mol Biol Evol 15:427-441. 81. Bravi CM, Bailliet G, Martinez-Marignac VL, Bianchi NO (2000) Origin of YAP+ lineages of the human Y-chromosome. Am J Phys Anthropol 112:149-158. 82. Weale ME, Shah T, Jones AL, Greenhalgh J, Wilson JF, Nymadawa P, Zeitlin D, Connell BA, Bradman N, Thomas MG (2003) Rare deep-rooting Y chromosome lineages in humans: lessons for phylogeography. Genetics 165:229-234. 83. Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (2008) New binary polymorphisms
85
FOCUS
reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res 18:830-838. 84. Thangaraj K, Chaubey G, Kivisild T, Reddy AG, Singh VK, Rasalkar AA, Singh L(2005)Reconstructing the Origin of Andaman Islanders. Science 308:996. 85. Palanichamy MG, Agrawal S, Yao YG, Kong QP, Sun C, Khan F, Chaudhuri TK, Zhang YP(2006)Comment on "Reconstructing the origin of Andaman islanders". Science 311:470. 86. Thangaraj K, Chaubey G, Kivisild T, Reddy AG, Singh VK, Rasalkar AA, Singh L (2006)Response to Comment on "Reconstructing the Origin of Andaman Islanders". Science 311:470. 87. 褚迅,单可人,文波,齐晓岚,李毅,吴昌学,刘烜,赵艳,任锡麟, 金力(2006)贵州瑶族3个支系Y-SNP及线粒体多态性研究. 遗传28:153-158. 88. 杜冠明(2008)汉藏语言的谱系.民族语文 (2):3-15. 89. Xue F, Wang Y, Xu S, Zhang F, Wen B, Wu X, Lu M, Deka R, Qian J, Jin L (2008) A spatial analysis of genetic structure of human populations in China reveals distinct difference between maternal and paternal lineages. Eur J Hum Genet 16:705-717. 90. Soares P, Trejaut JA, Loo JH, Hill C, Mormina M, Lee CL, Chen YM, Hudjashov G, Forster P, Macaulay V, Bulbeck D, Oppenheimer S, Lin M, Richards MB(2005) Climate change and postglacial human dispersals in southeast Asia. Mol Biol Evol 25:1209-1218. 91. 石泰安(著),耿升(译)(1992) 川甘青藏走廊古部落. 成都: 四川民族出版社. 92. 张亚莎(2007)古象雄的"鸟图腾"与西藏的"鸟葬".中国藏 学 (3):45-54. 93. 石硕(2001)藏族族源与藏东古文明.成都:四川人民出版 社. 94. 才让太(2006)苯教在吐蕃的初传及其与佛教的关系.中国
COM. on C. A. 2:e11, 2008
藏学 (2):237-244. 95. 赵心愚(2003)纳西族与藏族历史关系研究.博士学位论文, 四川大学. 96. 冯智(2008) 东巴教与滇西北苯教流行史迹试探. 中国藏 学(3):35-39. 97. 孙林(2006)试论苯教的宗教性质及与藏区民间宗教的关 系.西藏研究(4):30-35. 98. 魏征(唐) 隋书.卷八十三.列传第四十八.党项传. 99. 陈宗样,邓文峰(1991)白狼歌研究(一). 成都:四川人民出 版社.116. 100. 方国瑜(1944)么些民族考.民族学研究集刊 4: 19-44. 101. 李绍明(1981)康南石板墓族属初探——兼论纳西族的族 源.思想战线 (6):70-76. 102. 周伟洲(2002) 多弥史钩沉.民族研究 (5): 62-68+108. 103. 陈宗祥(1981)试论格萨尔与不弄(白兰)部落的关系.西南 民族学院学报(4):21-30. 104. 陈宗祥(2008)隋唐婢药(附国)历史研究——兼论该国为 《格萨尔王传》重要史料来源之一. 中国藏学(3): 28-34. 105. 娘毛加(2006)略谈《格萨尔》史诗中邓玛的身世及故乡. 硕士学位论文, 西北民族大学. 106. 谭其骧(1996)中国历史地图集. 北京:中国地图出版社. 107. 尊胜(2001)格萨尔史诗的源头及其历史内涵.西藏研究 (2): 31-44. 108. 赵秉理(2003)格萨尔王是人而不是神.青海民族学院学 报(社会科学版) (1):9-15. 109. 杨嘉铭(2008)关于英雄史诗主人公岭格萨尔王是否有 原型的讨论. 康定民族师范高等专科学校学报 (3):1-5. 110. 尕藏才旦(1991)岭尕尔是古代藏族建立在青藏高原上的 政治实体.西北民族大学学报(哲学社会科学版) (1):67-74. 111. 马岱川,扎西东珠(1987)岭尕尔与白兰—格萨尔"故乡" 初探.民族文学研究 (S1):21-31. 112. 才让(2007)吐蕃史稿. 兰州:甘肃人民出版社. 113. 陈立明(2003)藏门珞民族关系研究.博士学位论文,四川 大学.
原文翻译
兰海 译
最早定居东亚的现代人的 Y 染色体证据以及西藏人和日本人 的多重起源
(原文的图表并未复制到译文中.)
摘要:先前的东亚Y染色体系统发育关系表明,非洲起源的现代人最早定居在东南亚的大陆部分,然后大约在 25,000-30,000年前向北迁徙并遍布整个东亚.但是,东亚特有的Y染色体单倍群D-M174的分布却与这个图景 不同: 它仅高频分布于西藏, 日本和安达曼群岛. 在本文的研究中, 我们从东亚的73个人群中搜集了超过5,000 个男性样本,重建了单倍群D-M174的系统发育关系.我们的结果表明,D-M174是东亚地区现代人的极其古 老的支系,在南北人群观察到了深度的分化.我们认为D-M174起源于南方,然后在约60,000年前向北方扩散. 这一扩散早于东亚其他单倍群的扩散.新石器时代汉文化的扩张和末次冰盛期是导致目前D-M174在东亚的不 规则分布的关键因素. 西藏人和日本人是两个早期人群的混合, 即两个东亚特有的主要单倍群--单倍群O和D.
介绍
Y染色体Alu多态(YAP,即M1)定义了全 球Y染色体系统发育树上一个很深的分支 [1].单倍群D/E进一步分化为下游单倍群 DE*, D和E(原文图1).单倍群D/E的分布极 具地域性, 三个支系的分布都局限在某一地 理区域, 因此对追溯人类早期的历史非常有 意义(原文表1). 单倍群D/E中可能是最古老 的支系单倍群DE*仅在非洲的尼日利亚人 中有发现[2].这一发现支持现代人类起源 的"走出非洲"模式. 由M40/SRY4064和M96 定义的单倍群E(E-M40)也被认为起源于非 洲[3-6],而后约在20,000年前扩散到中东和 欧洲[3,4]. 有意思的是, 东亚特有的由M174 定义的单倍群D(D-M174)在西藏人和日本
人中比例很高(D-M174),但在其他东亚人 群以及邻近东亚(中亚,北亚和中东)的人群 却极少(通常少于5%) [5-7]. 日本人的D属于 D-M174下面的一个独立的单倍群,这个单 倍群由多个突变定义(例如M55, M57 和 M64等).这个分支与西藏人的不同,表明 它们之间是深度分化的[1].D-M174在东亚 的不规则分布与东亚的其他特有支系不同, 例如单倍群O下面的O3-M122,O1-M119和 O2-M95 [8, 9]. 在西藏和日本之外,D-M174 也普遍存 在于东亚南方的一些族群, 包括中国西南云 南省的操藏缅语的各个人群(14.0-72.3%), 中国南部广西省一个苗瑶语人群(30%)[ 编 者按:实指侗台语的拉珈人]和泰国的一个侗台
86
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 语人群(10%).这可能可以解释为很晚近的 人群混合[9-11].不过,最近的一项研究报 道 了 安 达 曼 人 中 有 高 频 的 D-M174(56.25%).他们是生活在印度洋中 偏远岛屿的人群, 被认为是东南亚最早的现 代人群体之一[12]. Underhill 等的另外一项 研究认为, D-M174 有可能在 50,000 年前到 达东亚[5].这表明东亚的 YAP 支系应该是 非常古老的. 我们之前的研究表明, 东亚特有的且在 东亚占主要的Y-SNP单倍群O-M175 (平均 44.3%)反映了现代人在东亚的早期的扩张 [8,9,13].与O-M175在大多数人群占主要的 情形不同的是,D-M174相对高频的人群主 要位于东亚大陆的外围区域,且分布离散 [7,9-11,13-18], 这可能意味着人类史前的两 次迁徙.一种观点认为,与O-M175一样, D-M174可能只是旧石器时代东亚人类向北 迁徙的多个单倍群之一[8,9]. 结果由于迁徙 之初人群本身的结构(末次冰盛期可能是关 键影响因素)以及后来汉文化[编者按:实指汉 文化的前身华夏文化]的扩张, D-M174的分布 被边沿化到现在的地理分布状态. 另外一种 推测是:D-M174的迁徙是一次独立的早期 迁徙,与我们之前提出的不同[8,9].为了回 答这个问题, 我们进行了全面的采样以及基 因分析, 分析涉及73个东亚和东南亚人群的 5,000个男性个体.基于Y-SNP/STR数据以 及D-M174主要支系的年代计算,我们认为 东亚地区的现代人在旧石器时代存在另一 次独立的向北的迁徙, 早于我们之前提出的 向北的迁徙[8, 9,13,19-21].
动态评论
M15, M57和P47的分型, 以便确定下游支系 的归属. 下游单倍群由相应的突变命名, 遵 从于YCC(2002)的规定.分型应用了PCR电 泳,PCR-RFLP和测序等方法[7].Y染色体 双等位标记的系统发育关系如原文图1所 示. 我们对5,134个样本进行了YAP基因座 的分型,结果测得512例YAP+(9.97%).加 上之前发表的207例Yap+样本,一共719例 Yap+进行了5个双等位标记(M174,M40, M15, M57和P47)和8个STR位点进行了检测 (DYS19/394, DYS388, DYS389Ⅰ, DYS389 Ⅱ , DYS390 , DYS391 , DYS392 和 DYS 393).719例Yap+样本中,697例得到了所 有SNPs和STRs的整组数值(数据见原文附 件1).
数据分析
在数据分析中包含了已经公开发表的 数 据 , 包 括 Hammer 等 (2006) 中 的 90 例 , Thangaraj等 (2003)中安达曼人的19例以及 Weale等(2003)中尼日利亚的6例.D-M174 下游支系的分化时间由STR数据计算, 采用 SNP-STR结合的方法[4,24,25].Y-STR的平 均突变速率采用0.00069[26].D-M174的各 个下游单倍群的Y-STR单倍型网络结构图 用 NETWORK4.2.0.1(www.fluxus-engineering.com) 来构建,然后分层次组合到已有的D-M174 系统发育树中(原文图1).各人群的平均遗 传基因多样性通过8个STR位点的等位基因 的 频 率 计 算 得 到 (Arlequin3.0, http://lgb.unige.ch/arlequin/).
方法
样本
本项研究中, 我们总共采集了73个人 群的5,134个无关男性样本.这些人群覆盖 了东亚和东南亚的绝大部分区域(原文表2 和图2).大部分样本来自中国南部和西南 部,中国80%的民族生活在这里超过3,000 年[22].之前报道的样本也包括:来自宿兵 等[7,9,17]的91个YAP+样本(日本16例,西 藏54例,朝鲜人3例,关岛人1例,柬埔寨人 1例, 泰国人4例以及来自中国其他地区的12 例)和来自Wells等[23]的116例 YAP+(原文 表2).
结果
原文表1列出了已经报道的世界范围内 人群的YAP+频率(根据参考文献的表注). 非洲有最高频的YAP+,全部属于下游单倍 群E-M40.而D-M174总体来说是一个亚洲 特有的, 并在邻近的地区有少量分布, 比如 中亚, 中东和东北印度. D-M174 在东亚的 平 均 频 率 为 9.60% , 高 频 出 现 在 西 藏 (41.31%) , 日 本 (35.08%) 和 安 达 曼 群 岛 (56.25%),而在东亚其他人群中较为罕见 (<5%).在进行Y染色体双等位基因标记 (M174,M40,M15,M57和P47)的分型之 后,719 例Yap+样本被划归6个下游单倍 群:DE*,E-M40,D*-M174,D1-M15, D2-M57 和 D3-P47 ( 见 原 文 图 1) . 719 例 Yap+样本的8个STR位点的进一步检测产 生了697组完整的数据. 如原文表2所示, 与 之前的报道一致[7,9-11,13,16],D-M174主
Y染色体标记和分型
首先对所有的样本都进行了3个Y染色 体双等位基因位点的分型, 包括M1, M40和 M174.属于单倍群D的样本进一步进行了
87
FOCUS
COM. on C. A. 2:e11, 2008
要高频存在于中国西部,南部和日本. D-M174的4个下游单倍群(原文图1)的 分布格局彼此不同.D1-M15在东亚广泛分 布,包括大多数藏缅语人群和侗台语人群 (原文表2).D*-M174和D3-P47主要分布在 藏缅语人群中,零星出现在侗台语人群中. 令人惊讶的是, 我们在西藏人中发现了两例 DE*,而这种类型之前仅在非洲被发现(尼 日利亚).与此不同的是,D2-M57仅出现在 日本,这暗示着这个支系很早就与D-M174 的其他下游单倍群分化开了(原文表2).我 们在西北汉族中识别出了4例E-M40,反映 了来自中亚的晚近的基因交流[23].为了得 到D-M174各支系的详细结构,我们结合 SNP和STR数据进行了网络结构分析(原文 图3).D*-M174有深层的结构,而且网络中 没有环路. D*-M174包含了藏缅语人群, 侗 台语人群和安达曼人的各自不同单倍型. 其 中没有观察到共享的单倍型(指的是不同地 区/不同语言的人群间),这表明D*-M174是 一个很古老的支系.作为一个最普遍的支 系,D1-M15的网络结构也呈现出深层的结 构, 通过很少的单倍型共享状态可以推知的 清晰的南北分化.与此不同,D2-M57局限 于日本,D3-P47在西藏高频出现而在邻近 的中亚和侗台语人群少量出现. 短距离和星 状的网络结构表明, 这两个单倍群在两个彼 此距离遥远的地区的长期存在和人口的扩 张.非藏族的藏缅语人群,像纳西族,普米 族和羌族的单倍型仅是藏族单倍型的子集. 这再一次证实了来自西藏的晚近的基因交 流,就像文献记载的那样[7, 22]. 接下来我们估计了D-M174各个下游支 系的年代.合并数据分析的结果表明, D*-M174, D3-P47和D1-M15的年代最为古 老,分别是66,392 ±1,466年,52,103±1,327 年和51,640 ±2,563年.日本特有的D2-M57 是最年轻的(37,678±2,216年) (原文表3).值 得注意的是, 得到的这些年代都比O3-M122 的年代古老得多. O3-M122是我们之前报道 另外一个东亚特有的单倍群(25,000-30,000 年) [8].
讨论
D-M174的不均匀分布可能导出另外一 种关于东亚早期人群的起源和迁徙历史的 推论.我们的数据不支持D-M174的北方起 源假说――因为中亚人群中的D-M174是罕 见的(原文表1),而且在网络结构图中中亚 的单倍型都处在外围的位置(原文图3).我
们的数据也不支持D-M174起源于印度的观 点――因为印度的996例样本中均没有检测 到D-M174. 而D-M174在印度东北的少量出 现(232例中有2例)是因为那些人群实际是 说藏缅语的人群(原文表1).西藏与印度之 间有限的基因交流可能是由于喜马拉雅山 脉的巨大的地理隔绝效果[27]. 另一方面,从Y染色体单倍型上看,居 住在安达曼群岛上的土著居民在基因上是 隔离的. 尽管在表型上与其他的东南亚人群 不同, 安达曼岛的居民拥有大部分东亚特有 的Y染色体支系,包括D-M174,O3-M122 和O2-M95.这一点强烈暗示他们是一个旧 石器时代人群的遗存[28].同样的,从语言 学和考古学证据看, 侗台语人群和苗瑶语也 是中国南方古老的人群[22].网络结构分析 显示D1-M15在南北人群(藏缅语人群对比 侗台语/苗瑶语人群)间有明显的分化(原文 图3).考虑到北亚没有D-M174以及D-M174 在中亚的零星出现,D-M174的北方起源假 说是不太可能的.因此我们可以确定 D-M174起源于南方, 这与之前提出的"最早 的现代人居住在东南亚的大陆部分"以及其 他Y染色体支系的迁徙图景是一致的[8, 9, 13]. 在多篇反对东亚人群的南方起源的文 献中, 相比南方人群, 在北方人群中观察到 了更高的遗传多样性[14,30]. 就像在我们之 前的报告中已经讨论的那样, 由于没有考虑 到来自中亚的晚近的混合,Karafet等[14]的 数据给出了一个北方的遗传多样性更高的 假象.尽管同时用到了Y-SNP和Y-STR数 据,薛雅丽等[30]的研究有同样的缺陷.在 薛雅丽等(2006)的文章中,蒙古人,维吾尔 人和满族人的遗传多样性更高, 而文献记录 中这些人群都与中亚人群或汉族有广泛的 混合[22].此外,薛雅丽等(2006)的文章中 南方的人群数较少, 长期地理上的隔绝导致 的瓶颈效应会极大地影响到测得的遗传多 样性. 当大量的南方人群被包含到研究中来 时, 我们观察到这些人群的遗传多样性比北 方人群的更高[8, 9]. 基于STR数据得到的南方人群的遗传 多样性与北方人群的相当. 西藏人有最高的 多样性(0.525±0.294),接下来是侗台语人群 (0.484±0.272), 日 本 人 (0.419±0.239) 和 苗 瑶 语人群(0.347±0.206).没有计算其他东亚人 群的遗传多样性是因为样本量太少. 西藏人 有最高的多样性主要是因为相比其他人群 西藏人有庞大的D-M174有效群体数量.西 藏人和日本人生活在两个彼此距离遥远的
88
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 地区,他们的D-M174分属于两个不同的下 游支系. 这两个单倍群都有短距离和星状的 网络结构, 这表明在同一地区的长期存在和 晚近的人口扩张(原文图3).需要注意的是, 遗传多样性的计算可能包含潜在的偏差, 比 如D-M174各支系年代的差异.之前仅在非 洲发现的DE*现在在西藏也有发现, 这支持 D-M174的古老程度以及D-M174存在于最 早定居在东亚的现代人之间. 此外, D-M174 的离散分布和它的古老年代支持一次旧石 器时代东亚现代人的迁徙.我们之前关于 O3-M122的数据显示了东亚地区现代人一 次史前(约25,000-30,000年前)向北的迁徙. 这种观点可以解释大部分现在东亚特有的 Y染色体支系的系统发育关系(O3-M122, O2-M95和O1-M119) [8,9,13].不过这个推 测的迁徙图景无法解释D-M174的分布.首 先,D-M174在东亚大陆的中心地带非常罕 见, 特别是在汉族人群中. 尽管这可以假设 为基因漂变的影响:假设D-M174在我们提 出的这次向北的迁徙中伴随着O3-M122 , 则D-M174在西藏人和日本人中的高频需要 重复发生的突变或独立且随机的富集过程. 这显然是不太可能的. 一次更早的独立的向 北迁徙可以合理地解释D-M174目前在东亚 的分布状态. 由于存在更晚的O3-M122的向 北迁徙以及新石器时代汉文化的扩张, 我们 推测D-M174在东亚中心地带的迁徙痕迹已 经被O3-M122更晚但也更大规模的迁徙覆 盖了.目前D-M174分布与东亚外围区域的 状态与这一假设符合. 同时, 计算得到的年 代也表明D-M174的迁徙早于O3-M122. 人们认为晚更新世早期(13.5至7.5万年 前)东非的大干旱强迫现代人走出了非洲 [31].早期的人类可能占据了沿海地区并且 将近水域的海产品作为食物[32].然后,现 代人沿海岸线扩散. 在非洲之外, 最早的现 代人脚印化石的年代约为10万年左右[33]. 8万至1万年间的末次冰期可能对现代人的 迁徙产生了巨大的影响, 当时的海平面在现 在的海岸线之下50-200米[34].这使更宽的 陆地浮现出来了,比如日本与亚洲大陆之 间, 使人们能够在现在被大海隔离的陆地间 迁徙. 人类化石记录以及之前的基因数据表 明,定居于东亚最早的现代人稍晚于6万年 [8,9,13,21, 35].比如,最早的澳大利亚古人 类化石(蒙戈湖3号坑)的年代在 45,000±3,000-62,000±6,000年之间[36, 37]. 而对现在的澳洲土著进行的mtDNA和Y染 色体数据分析表明人类约在50,000-70,000
89
动态评论
年 前 出 现 在 澳 大 利 亚 [38] . 我 们 得 到 的 D-M174的年代与以上观点是相符的,尽管 现代人进入东亚和澳大利亚可能是彼此独 立的迁徙事件[38].计算得到的D-M174的 年代比之前报道的基于Y染色体和mtDNA 数据的年代更加古老[8, 9, 21].为了确定年 代是否被过高估计, 我们使用同样的方法计 算了DE*与E-M40的分化时间.计算得到的 年代是27,176年,比D-M174的年代年轻得 多,但与之前的(约27,800-37,000年)报道接 近[3].因此,D-M174的古老确实反映了东 亚早期人类的迁徙. Zhivotovsky (2001)提出 的年代计算方法对有效人口数量以及晚近 的人口扩张不敏感, 尽管人群本身的结构也 不能完全忽略.D-M174的古老也得到之前 的研究的支持,在那篇文献中D-M174的年 代超过50,000年[5]. 单倍群D的分化年代约为6万年,这与 它现在宽广但离散的地理分布是一致的. 我 们提出的旧石器时代的迁徙可能是现代人 定居东南亚之后向北的最早的一次迁徙. 因 为末次冰期发生在8万至1万年前之间, D-M174向北的迁徙与之前提出的观点相 符――人类可能以"猛犸大草原"上的食物 为生[39].除了后来的人口扩张,末次冰期 可能也对D-M174现在离散的分布产生了影 响. 值得注意的是, 最近的一项考古研究发 现, 人类可能在30,000-40,000年前就来到了 青藏高原[40].这比之前认为的要早的多, 但是与我们的结果相符. 冰后期海平面的上 升最终导致了日本与亚洲大陆的分离, 这可 以解释D-M174现在在日本人群中的遗存. 考古学数据证实现代人约在30,000年前开 始在日本定居[41, 42],与我们计算得到的 D2-M57的年代(约37,678 ± 2,216年)相符. 总而言之,目前西藏人和日本人应是由 D-M174和O3-M122分别代表的两个古老人 群的混合[7, 10, 16].
结论
总结认为, 我们论证了东亚人群旧石器 时代的一次迁徙. 这次迁徙早于之前提出的 向北的迁徙.现在D-M174的离散分布主要 归因于新石器时代晚期的人群扩张和末次 冰期.
原文参考文献
1. Y-Chromosome-Consortium(2002)Genome Res 12:339-348. 2. Weale ME et al.(2003)Genetics 165:229-234. 3. Cruciani F et al.(2007)Mol Biol Evol 24:1300-1311. 4. Semino O et al.(2004)Am J Hum Genet 74:1023-1034. 5. Underhill PA et al.(2001)Ann Hum Genet 65(Pt 1):43-62. 6. Underhill PA et al.(2000)Nat Genet 26:358-361.
FOCUS
7. Su B et al.(2000)Hum Genet 107:582-590. 8. Shi H et al.(2005)Am J Hum Genet 77:408-419. 9. Su B et al.(1999)Am J Hum Genet 65:1718-1724. 10. Hammer MF et al.(2006)J Hum Genet 51:47-58. 11. Wen B et al.(2004)Am J Hum Genet 74:856-865. 12. Thangaraj K et al.(2005)Science 2005 308:996. 13. Jin L&Su B (2000)Nat Rev Genet 1:126-133. 14. Karafet T et al.(2001)Am J Hum Genet 69:615-628. 15. Ke Y et al.(2001) Science 292:1151-1153. 16. Qian Y et al.(2000)Hum Genet 106:453-454. 17. Su B et al.(2000)Proc Natl Acad Sci U S A 97:8225-8228. 18. Wen B et al.(2004)Nature 431:302-305. 19. Ballinger SW et al.(1992) Genetics 130:139-152. 20. Chu JY et al.(1998)Proc Natl Acad Sci U S A 95:11763-11768. 21. Yao YG et al.(2002)Am J Hum Genet 70(3):635-651. 22. Wang ZH(1994) History of nationalities in China. Beijin: China Social Science Press. 23. Wells RS et al.(2001)Proc Natl Acad Sci U S A 98:10244-10249. 24. Rootsi S et al.(2004)Am J Hum Genet 75:128-137. 25. Zhivotovsky LA et al.(2001) Mol Biol Evol 18:700-709. 26. Zhivotovsky LA et al.(2004)Am J Hum Genet 74:50-61. 27. Cordaux R et al.(2004)Mol Biol Evol 21:1525-1533. 28. Thangaraj K et al.(2003)Curr Biol 13:86-93. 29. Lell JT et al.(2002)Am J Hum Genet 70:192-206.
COM. on C. A. 2:e11, 2008
30. Xue Y et al.(2006)Genetics 172:2431-2439. 31. Scholz CA et al.(2007)Proc Natl Acad Sci U S A 104:16416-16421. 32. Walter RC et al.(2000)Nature 405:65-69. 33. Forster P&Matsumura S(2005) Science 308:965-966. 34. Lambeck K et al.(2001)Science 292:679-686. 35. Wu HC et al. (1995) Human Evolution in China: A Metric Description of the Fossils and a Review of the Sites. Oxford: Oxford Univ Press. 36. Bowler JM et al.(2003)Nature 421:837-840. 37. Thorne A et al.(1999)J Hum Evol 36:591-612. 38. Hudjashov G et al.(2007)Proc Natl Acad Sci U S A 104:8726-8730. 39. Guthrie RD(1990)Frozen fauna of the Mammoth Steppe. Chicago: University of Chicago Press. 40. Yan YB et al.(2007)Chin Sci Bull (Chin) 52:1567-1571. 41. Ono A et al.(2002)Radiocarbon 44:477-494. 42. Takamiya H et al.(2002)Radiocarbon 44:495-502. 43. Cinnioglu C et al.(2004)Hum Genet 114:127-148. 44. Cruciani F et al.(2002) Am J Hum Genet 70:1197-1214. 45. Hammer MF et al.(1998)Mol Biol Evol 15:427-441. 46. Jin HJ et al.(2003)Hum Genet 114:27-35. 47. Kivisild T et al.(2003)Am J Hum Genet 72:313-332. 48. Ramana GV et al.(2001)Eur J Hum Genet 9:695-700. 49. Semino O et al.(2002)Science 290:1155-1159. 50. Zerjal T et al.(2002)Am J Hum Genet 71:466-482.
原文翻译
兰海 译
YAP 在南亚的分布状态
(原文的图表并未复制到译文中.)
介绍
人类Y染色体Alu多态(YAP)单元是研 究人类进化的热点,因为它位于人类Y染色 体的非重组区(Hammer 1994).Y染色体中携 带YAP插入元件的频率在不同地区的人群 差异极大(Hammer 1994;Hammer & Horai 1995; Hammer et al. 1997; Karafet et al. 1997). YAP插入 多态在某些亚洲群体, 太平洋群体以及印第 安群体低频(<10%)存在,在西部欧亚群体 中频率适中(11-25%), 而在以下群体的频率 超过30%:日本,藏族以及多个非洲人群 (Hammer 1994; Spurdle et al. 1994; Hammer &
Horai 1995; Altheide & Hammer 1997;Bianchi et al. 1997; Karafet et al. 1997; Hammer et al. 1998; Thomas et al. 1998; Cruciani et al. 2002).YAP插
材料与方法
我们在印度不同地区的21个人群中收 集 了 总 共 2169 例 血 样 , 然 后 根 据 Hammer&Horai (1995) 的方法检测了YAP 插入状态. 属于YAP+的样本进一步用M174 和M96的引物进行了分型, 以便确定具体属 于单倍群D或E.
结果与讨论
2169例样本中, YAP插入出现在73例样 本中(见原文表1). 其中只有古吉拉特Dungri Bhill的1例YAP样本属于单倍群E,可以推 测是来自非洲或中东人群的晚近混合. 被认 为起源于中东的北印度穆斯林人群Shia人 中有11%的E(Agrawal et al. 2005).因含有 M174突变而归属于单倍群D的72例YAP插 入 多 态 样 本 包 含 Dirang Monpa (34%) , Shertukpen (8%),Lepcha (16%), Lachungpa (65%) 和 Jarawa (100%) 部 落 . 我 们 对 Jarawa检测的结果与Thangaraj等(2003)的发 现一致. 我们已经确定安达曼群岛的三个部 落(昂格人,大安达曼人和贾拉瓦人)是现 存的尼格利陀人,他们分享共同的文化背 景, 语言以及mtDNA单倍群和Y染色体单倍 群D*.他们与亚洲人群而不是非洲人群更 接近(Thangaraj et al. 2003, 2005; Palanichamy et al. 2006) . 属 于 D* 的 单 倍 型 在 中 亚 有 发 现 (Karafet et al. 2001).YAP下的单倍群D与E的
入多态在南部非洲的极高频率分布使得人 们认为YAP起源于非洲然后扩散到别的大 陆. 不过,Hammer等(1997, 1998)和Altheide &Hammer (1997)提出YAP亚洲起源的另一 种假说. 我们已经知道YAP插入多态分成两 大支系:单被群E分布在非洲以及地中海沿 岸,单倍群D分布于日本以及一些东南亚群 体.印度的Y数据显示,现在印度人群的父 系主要还是继承自最早的居民而不是晚近 的融合(Cordaux et al. 2004; Sengupta et al. 2006). 为了说明人群进化的历史,我们检测了21 个印度人群中的YAP多态.
90
兰海:Y 染色体单倍群 D 在东亚的分布及其意义 系统发生关系尚不确定. 在尼日利亚发现了 5例超单倍群样本DE*使得Weale等(2003)能 够更仔细地研究YAP内部分支的顺序,认为 YAP支系的起源尚无法得到确定的. 暂不考 虑DE*,D*和E*的分支顺序,"安达曼岛居 民是来自亚洲大陆的移民"观点值得进一步 研究.
动态评论
进化学意义
非洲之外的人群都属于M168. M168分 成三个支系:YAP(DE),C (RPS4Y/M216) 和F* (M89/M213). Underhill等(2001)认为含 有M168突变的一个非洲人群从非洲之角开 始扩散,经过海岸或内部路线在5万年前到 达 南 亚 (Walter et al. 2000) , 单 倍 群 C(RPS4Y/M216)可能就诞生在那里.YAP 可能诞生在一个55000年前的亚洲Y染色体 上(Hammer et al. 1998).证据是在亚洲特有的 M174上有M40和M96的等位基因的祖先型 (Altheide & Hammer 1997). 但是, M174的祖先 型仅在非洲有发现, 支持YAP插入的非洲起 源. 但是突变事件发生的年代表明亚洲YAP 插入Y染色体上的的M174更为古老.我们 的数据显示, YAP出现在印度东北的部落以 及安达曼人(单倍群D)中.这表明南亚的 M168诞生了YAP插入以及M174突变.C*, YAP插入和F*在印度的出现表明南亚的Y 染色体已经分化成主要的下游支系(Kivisild
et al.2003; Cordaux et al. 2004; Sengupta et al. 2006; Thangaraj et al. 2003).然后他们迁徙到东南亚
M168诞生的主要的Y染色体下游支系并不 是非洲起源的.同样,没有迹象表mtDNA 超单倍群M,N和R是非洲起源的.根据 Olivieri et al. (2006)的研究,有两个mtDNA 单倍群的分布支持向非洲的回迁:M1(分子 钟计算的年代为3.86±0.71万年)和U6(分子 钟计算的年代为4.51±0.69万年),这两个诞 生在亚洲西南的支系是北非的主要mtDNA 单倍群, 它们在地中海沿岸区域分化出主要 的下游支系,不久之后M1a (分子钟计算的 年代为2.88±0.49万年),U6a2 (分子钟计算 的年代为2.40±0.73万年)和U6d (分子钟计 算的年代为2.06±0.73万年)通过黎凡特扩散 到东非和北非. 因此现代人将南部的海岸线 作为"走出非洲"的出口. 总之,单倍群 D 显示出非常不同的空 Jarawas 人中 100%的 YAP(单倍群 间分布. D)是由于人所共知的人口统计学历史(长期 的基因隔离)引起的奠基者效应(Thangaraj et al. 2003).YAP 在某些东北印度部落中高频 出现可能是由于基因漂变.此外,YAP 在 别的印度人群中没有发现, 除了南部印度卡 纳塔克邦的 Koraga 部落的少量发现(0.6%, 2/33,Cordaux et al. 2004).首次走出非洲的迁 徙中 M168 采用的应该是南部路线.YAP 插入单倍型的这个支系表明安达曼人可能 是最古老的人群的后裔.根据 Olivieri et al. (2006)的发现, 东北印度部落中 YAP 的发现 对理解区域进化历史有重要的意义.
原文参考文献
Agrawal S et al.(2005)Curr Sci 88:1977-1980. Altheide TK et al.(1997)Am JHum Genet 61:462-466. Bianchi NO et al.(1997) Am J Phys Anthropol 102:79-89. Cordaux R et al.(2004)Curr Biol 14:231-235. Cruciani F et al.(2002)Am J Hum Genet 70:1197-1214. Cruciani F et al.(2004)Am J Hum Genet 74:1014-1022. Hammer MF et al.(1994)Mol Biol Evol 11:749-761. Hammer MF et al.(1995)Am J Hum Genet 56:951-962. Hammer MF et al.(1997) Genetics 145:787-805. Hammer MF et al.(1998)Mol Biol Evol 15:427-441. Hammer MF et al.(2006)J Hum Genet 51:47-58. Karafet TM et al.(1997)Am J Phys Anthropol 102:301-314. Karafet TM et al.(2001) Am J Hum Genet 69:615-618. Kivisild T et al.(2003)Am J Hum Genet 72:313-332. Luis JR et al.(2004)Am J Hum Genet 74:532-544. Olivieri A et al.(2006)Science 314:1767-1770. Palanichamy MG et al.(2006)Science 311:470a. Sengupta S et al.(2006)Am J Hum Genet 78:202-221. Spurdle AB et al.(1994)Am J Hum Genet 54:319-330. Su B et al.(1999)Am J Hum Genet 65:1718-1724. Thangaraj K et al.(2003)Curr Biol 13:86-93. Thangaraj K et al.(2005)Science 308:996. Thomas MG et al.(1998)Nature 394:138-140. Underhill PA et al.(2000)Nat Genet 26:358-361. Underhill PA et al.(2001)Ann Hum Genet 65:43-62. Walter RC et al.(2000)Nature 405:65-69. Weale ME et al.(2003)Genetics 165:229-234.
和安达曼群岛. 安达曼人的母系支系与西孟 加 拉 邦 的 拉 吉 班 什 人 (Rajbansi of West Bengal)(Palanichamy et al. 2006)和奥里萨邦的 保里布伊亚人(Pauri Bhuiya of Orissa,数据 未公布)的mtDNA单倍群M31建立了联系. 在到达东亚南部之后, 最初扩散的人群的后 代 向 北 扩 散 最 终 遍 布 整 个 东 亚 (Su et al. 1999).而其他的没有携带M174突变的YAP 插入染色体向西通过西亚进入地中海沿岸, 诞生了带有M40和M96突变的单倍群E (约 31,000年前; Hammer et al. 1998). Hammer等(1997)和Altheide & Hammer (1997)提出假说, 认为单倍群E通过黎凡特向 非洲回迁.有证据表明,E-M34就是由近东 进入埃塞俄比亚的(Cruciani et al. 2004).从亚 洲回迁的假说得到Y染色体变异的系统地 理分布的支持: 单倍群K2以及R1b*属于亚 洲的超单倍群K,却仅在非洲的高频出现 (Cruciani et al. 2002; Luis et al. 2004).



