中央研究院 植物研究所 杜 镇 研究员
一,质粒
绝大多数的生物都是以DNA的形式来储藏其遗传信息.遗传物质要能生生不息地传给後代的首要条件就是它至少要具有一个复制原(ori,origin ofreplication,或译为复制起点),使整个基因体得以复制.含有复制原的遗传物质称为replicon,我们姑且把它译为为复制体吧!.原核性复制体分为原核染色体,质粒(plasmids)和噬菌体基因体(phagegenome)等三类.其中质粒的基因体和原核染色体类似,是由双绞鍊DNA构成,并以超卷曲的形式存在.它们的基因体约由2,000至150,000个碱基对组成,绝大多数呈环状,但也有极少数是线状构造(如Borreliaburgdorfferi).事实上你可以把它们视为比较小的原核染色体.在自然环境中它们相当普遍地生存在原核生物细胞内,并和其宿主的许多特殊功能有关,诸如:赤贺氏杆菌(Shigella)的抗药,根瘤菌(Rhizobium)的固氮,农杆菌(Agrobacterium)的引瘤及假单胞杆菌(Pseudomonas)对环状有机物的分解等等.以下我们谈的以细菌性质粒为主,尤其是革兰氏阴性菌的质粒.
二,质粒的类型
当我们谈到质粒的类型时,就要看你从哪个角度来看它们,譬如说抗药性,结合生殖能力,宿主范围及DNA复制方式等等.这些分型标准之间并无横向关联.你无法说能结合生殖的质粒一定抗药或不抗药,也无法确定宿主范围和质粒套数的调控有何关联.我们用到这些名词时,只是对特定质粒的性状做一些描述而已.质粒的真正系统分类标准并非靠些性状,而是依据它们的不共容性(incompatibility).
有的质粒带有显著特徵可供我们侦测它们的存在,无已知特徵的质粒称为隐性质粒(crypticplasmids);有特徵者称为显性质粒(acrypticplasmids);带有抗药基因的天然质粒称为R-质粒(R-plasmids).有些质粒能在多种不同菌属细胞中生存,我们称它们为泛宿主性质粒(broad-host-rangeplasmids);有一些质粒只能在少数相关宿主中生存,我们称它们为狭宿主性质粒(narrow-host-rangeplasmids).具有结合生殖(conjugation)能力的质粒为结合质粒(conjugativeplasmids);没有这种能力的质粒便是非结合质粒(non-conjugativeplasmids).其他如侵袭性质粒(virulence plasmid),共生质粒(symbioticplasmids)及巨型质粒(megaplasmids)等等有关质粒性状叙述的名词不一而足.
三,质粒的复制
环状质粒的复制形式主要分theta (θ)及rollingcircle两种.基因体的复制都是由复制原开始.原核性复制原约由250个碱基对组成,一般质粒的复制原常称为oriV(origin of vegetativereplication);有时R-质粒的复制原称为oriR;大肠杆菌的复制原称为oriC.Theta形式的质粒复制与细菌基因体复制一样,以RNA聚合脢(RNApolymerase)在复制原制造RNA引子(RNA primer),然後由DNA聚合脢(DNApolymerase)接手由此向两个方向分别复制DNA,直到整个基因体复制完成.在复制过程中当然还有许多其它酵素的参与,这些酵素多由宿主提供.有的质粒自己携带一些与复制有关的基因,这些基因多被命名为rep,如repA,repB等等.Rollingcircle形式的质粒复制与部分噬菌体基因体的复制相似.环状双绞链中的一股在复制原附近被酵素切开,然後环状股与线状股被分别复制完成.至於线状质粒的复制则是靠特殊的RNA引子或该质粒所具有的telomere-likesequence.
四,质粒的套数与套数调控(copy number control)
在一个宿主细胞中某一特定质粒出现的个数称为套数copynumber.在大肠杆菌中,质粒的套数由F-质粒(fertilityplasmid)的一个到pUCs的数百个不等.质粒套数的控制与该质粒复制的调控机制有关,几乎每一种质粒都有它独特之处.有关质粒套数的控制我们仅以两种代表型机制来说明.
1. 反意RNAantisense RNA:
ColE1质粒的复制是由RNA聚合脢由其复制原(oriV)前方555个碱基对以前的启动子(promoter)读出一段称为RNAII的产物.这个称为前引子(preprimer)的RNAII会利用碱基配对而自我摺叠成可被RNAaseH(一种特殊的RNA水解脢)裁剪的立体结构.RNAII经适当的裁剪後就形成正确的复制引子,复制於焉开始.在ColE1质粒复制原前方445个碱基对的後方另有一个反向的启动子.RNA聚合脢由此处转录出一条称为RNAI的产物,它与RNA II反向,刚好形成配对,这种RNA称为反意RNA(antisenseRNA).当RNA I和RNA II形成RNA-RNA杂合体(RNA-RNA hybrid)时,RNAII无法形成可被RNAaseH剪裁的结构,复制引子无法产生,复制也不会有效地开始.当然,实际上还有许多辅助蛋白和酵素会控制RNAI和RNA II之间的亲合力和RNA I的水解速度.RNA I和RNAII的相对数量的增减决定了ColE1质粒的复制速度,也决定了该质粒的套数.
2. Iterons与repA:
质粒pSC101的复制除了需要宿主供应的酵素外尚需要RepA蛋白.RepA是pSC101的基因产物,在功能上他扮演了复制的活化因子(activator)与自我抑制因子(autorepressor).RepA基因位於pSC101复制原的前方,其转录方向和pSC101复制方向正好相反.在RepA基因的启动子和pSC101复制原中有好几个顺向和反向的重复DNA短段,长约18~22个碱基对,称为iterons.RepA会粘黏在iteron上.质粒在低浓度(低套数)时,产生低浓度的RepA促进了质粒的复制;质粒在高浓度(高套数)时,产生大量的RepA粘黏在复制原和RepA基因的启动子上抑制质粒的复制.於是,RepA和iterons的相互作用决定了pSC101的套数.
五,质粒的不共容性(incompatibility)
有时我们会想知道一个细菌细胞中会同时出现多少种质粒呢(种类非个数,个数是指套数)这个问题倒没有一个确切的答案.由自然环境分离的细菌细胞中含有一至三种质粒是常见的,有时多至七,八种也不令人意外.质粒的种类虽多,但并非所有的质粒都能同时在同一个宿主细胞中生存.有一些质粒在同一个宿主细胞中会发生相互排斥的现象,我们称这种现象为质粒的不共容性(incompatibility).质粒的不共容与他们的复制机制与套数调控有关.当两种复制与套数调控机制毫不相干的质粒在同一个细胞中生存时,它们各自复制再各自分配到不同的子细胞中,不会发生干扰.当两种复制机制与套数调控相关或甚至相同的质粒在同一个细胞中生存时,复制与套数调控机制就会把它们看做同一种质粒而将他们的套数控制在一定的数目内.因为细胞分裂时质粒在子细胞中分配不均匀的缘故,经过几十次的细胞分裂後你只可能在特定的子细胞中找到其中一种质粒.根据这种不共容性,质粒被分为许多不共容群(incompatibilitygroup,简称为inc),譬如RP4属於incP;RSF1010属於incQ而pSa属於incW等.如果你在实验中发现有一种质粒竟然属於两个不同的不共容群或有不共容不完全的现象.别惊慌!那是有可能发生的.在革兰氏阴性菌中,大肠杆菌与假单胞杆菌的质粒分类是做的比较清楚的.
六,质粒的宿主范围(host range)
质粒间的宿主范围有很大的差异.能在多种不同菌属的细胞中生存的质粒称为泛宿主性质粒,如RP4和RSF1010;只能在单一或极为相近的宿主中生存的质粒称为狭宿主性质粒,如F-质粒和ColE1.质粒的宿主范围与其复制所需要的条件和质粒本身所携带的基因有关.如果一个质粒携有与本身复制有关的基因,而这些基因的启动子又是最容易启动的,那它对宿主的需求自然很少,它的宿主范围当然很广.如RP4在几乎所有的革兰氏阴性菌中都能生存.相反的,一个对宿主需求很多的质粒,对宿主的依赖自然很强,它的宿主范围当然很窄.如F-质粒只能在大肠杆菌中生存.
七,质粒的结合生殖(conjugation)
有的质粒具有在细菌细胞间进行结合生殖的能力,这些质粒称为结合质粒(conjugativeplasmids)或自动转移质粒(self-transmissibleplasmids).质粒的结合生殖过程包含了细菌细胞间的结合与质粒的DNA复制.与质粒结合生殖有关的基因很多,其中包括主持其事的基因和许多调控结合生殖的基因.这些基因在不同的质粒系统中各有其名,但多数被命名为tra(Transfer)或mob(mobility),如traA,tra,mobA,mobB等等.一个结合质粒至少要具有下列三个基本条件:1.产生性线毛(sexpili)的能力,2.具有转移原oriT(origin oftransfer)及3.能辨认oriT的特殊DNA内切脢(DNA endonuclease).
结合质粒有产生线毛蛋白(pilin)的基因,线毛蛋白产生後组成性线毛.虽然性线毛因来源质粒不同而有F-线毛(来自F-质粒),R-线毛(来自R-质粒)等之分,但在功能上它们的作用都是用来刺激细菌细胞使之结合.含有结合质粒能产生性线毛的细菌细胞称为雄型(maletype)或正型(+ type);不含结合质粒无性线毛的细菌细胞称为雌型(femaletype)或负型(-type).结合生殖时正型称为授株(donor);负型称为受株(recipient).结合生殖的结果使负型变为新的正型,此时新的正型称为transconjugant(有些情况称为exoconjugant).
质粒在结合生殖时要靠特殊的DNA内切脢将质粒DNA中的一股在特定的地方切成单链,然後将切成单链的一股DNA经由两个细胞的结合处传送到受株中,同时以rollingcircle方式完成各股DNA的复制.质粒DNA中被切开的特定的地方就是oriT或称为bom(basisof mobility),nic(nicksite),它们具有一定的DNA序列.切oriT的DNA内切脢有时称为nickase,它是tra基因中之一的产物.每一类的oriT均有它特定的DNA内切脢,并且内切脢应该切oriT处的哪一条链及哪两个碱基之间也是一定的.
非结合质粒中有些因为具有oriT或bom,所以可以经由具tra基因的helper质粒协助而被传送.可被传送的非结合质粒称为可移动质粒(mobilizableplasmids),pBR322就属於这个类型;不可被传送者称为不可移动质粒(non-mobilizableplasmids),RSF1010属这一型.可移动质粒与helper质粒的搭配使用在基因转殖上是常被利用的技术.
结合质粒F上有若干个与大肠杆菌染色体上相同的转位子.在偶然的状况下因为相同DNA序列的转位子发生同源重组(homologousrecombination),结果使F-质粒与染色体DNA融成一体.这种授株被称为Hfr(highfrequency ofrecombination).当Hfr的F-质粒做结合生殖时,整个与其黏合的大肠杆菌染色体会依序地被传送到受株中.大肠杆菌的基因图谱就是靠这种方式完成的.你也可以利用泛宿主性结合质粒在其它菌种中做类似的工作,但因DNA定序技术的高度发展已使得这类工作显得费时与不够精确了.
八,载体(vector)
载体的主要功能在基因选殖(cloning),选殖的主要目的是分离,纯化及繁殖特定的基因.细菌性载体主要有质粒载体(plasmidvector)和噬菌体载体(phagevector)两种,我们这里所谈的以质粒载体为主.质粒载体的基本条件是:1.它们是复制体,能生生不息地被复制;2.它们有适当的标示基因,可供做转形株(transformant)和非转形株(non-transformant)的辨识;3.它们有另一组具有限制脢单切点的标示基因,可供插入外来基因并做为选殖株(clone)和非选殖株(non-clone)的辨识.此外它们的基因体要小,以便於操作和细胞转形.
利用质粒发展成载体第一个最成功的例子是pBR322(Bolivar et al,1977).它含有pMB1的复制原,pSC101的抗四环黴素(tetracycline)基因tet和Tn3(一种转位子)的抗青黴素(penicillin)基因bla.你可以把外来基因插在这两个抗药基因中的任何一个内,利用中断抗药基因的消失做选殖筛选,利用剩下的一个完整抗药基因做转形株筛选.因为pBR322的成功构筑,才使基因选殖有了长足的进展.
细胞转形(celltransformaiton)是基因选殖的重点工作之一.但细胞的转形率与外来DNA的大小成级数反比.以几万碱基对大小的DNA来转形大肠杆菌细胞几乎是不可能的.为了改善这种情况,结合了pBR322的载体特性和噬菌体λ(lambda)的壳蛋白包裹(packing)特性发展出cosmidpHC79 (Hohn and Collins1980).噬菌体λ的基因体由48,502个碱基对构成.它在宿主中的DNA复制产物是多个λ基因体相连的多元体(concatemers).当噬菌体壳蛋白接近完成时才将λ基因多元体由一端「吸入」.当壳蛋白已经包裹了约相当於λ基因体75%的DNA(约36,000碱基对)时,一个特殊的λDNA内切脢开始被活化并寻找适当的切点以便切开λ基因多元体完成包裹动作.如果壳蛋白已经包裹了约相当於λ基因体110%的DNA(约54,000碱基对)还没发现适当的切点时,包裹动作就会被放弃.这个「适当的切点」平时就在线状λ基因体DNA的两端,称为cohesive sites(cos).将噬菌体λ的cos切出植入pBR322中使新构筑的载体兼有质粒载体与可被视为λ基因体而被包入λ壳蛋白中的特性,可使30,000~50,000碱基对大小的DNA轻易地被选殖出来.因为它结合了λcos site和plasmid,故称为cosmid.
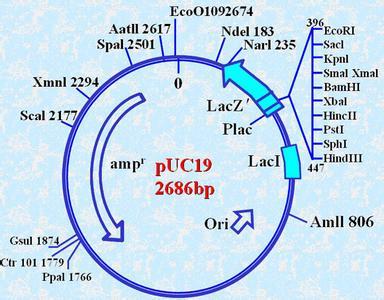
载体pUCs(如pUC18,pUC19,Yanisch-Perron et al,1980)则是利用pBR322做了选殖株筛选的进一步发展.它的特点是利用了呈色基因取代了pBR322中的一个抗药基因.大肠杆菌乳醣分解脢(β-galactosidase)可分解许多β-半乳醣化合物(β-galactoside),其中也包括了X-gal(5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside).X-gal呈淡黄色,分解後呈蓝色.大肠杆菌的乳醣分解脢有两个不寻常的特色:1.该酵素N-端的廿几个氨基酸和酵素活性无关,可以任意修改成连串的限制脢切点MCS(multiple cloning site)而不影响酵素活性;2..该酵素基因如果被适当地分成两段(如前段lacZ'和後段lacZΔM15),这两段基因的产物(α-chain和β-chain)尚能互补而具有酵素活性.载体pUCs利用了pBR322的基本骨架,以突变除去抗青黴素基因中的PstI切点,切除抗四环黴素基因并在该处加入具有MCS的lacZ'.至於後段的lacZΔM15则是利用F-质粒或噬菌体带入乳醣分解脢缺失型的大肠杆菌中.利用了这种载体与宿主的基因互补,pUCs改善了选殖株筛选的方式.何况在培养基中加乳醣分解脢基因的诱发物(inducer)IPTG(isopropyl-1-thio-β-D-galactopyranoside),更可诱发乳醣分解脢大量表现.你只要看颜色就能鉴别选殖株与非选殖株.
载体pBluescripts(如pBluescript II KS(+),pBluescript IIKS(-),(Alting-Mees and Short,1989))则是利用pUCs做了DNA不同形式生产的进一步发展.它的特点是结合了pUCs的质粒载体特色与线状噬菌体的DNA复制特色,故称为phagemid.线状噬菌体(filamentousphage)是一群以线状单链DNA为基因体,以大量单一种类蛋白质(B蛋白)呈螺旋状包裹的极细长噬菌体.它们藉由性线毛的协助感染宿主,基因体进入宿主後会环状化(circulization)并复制互补链成为双绞链.DNA复制以rollingcircle方式产生大量单链基因体DNA.基因体DNA被壳蛋白包裹後从宿主中「穿出」,并不会杀死宿主.将线状噬菌体f1的复制原切出植入质粒中并不会影响质粒的复制.但如果以适当的helper线状噬菌体感染含有phagemid的宿主时,phagemid就会产生大量的单链DNA.这些phagemid单链DNA也会被包入噬菌体壳蛋白中,并且包入的单链DNA与当初f1复制原在phagemid上的方向有关(+或-).如此你就可以从宿主细胞中纯化phagemid双链DNA从事选殖等用途;从噬菌体中纯化单链DNA从事定序工作.
载体pETs(如pET12a,pET25b,Moffatt and Studier,1986)则是结合了pBR322,pUCs或pBluescripts等的载体功能与噬菌体T7的基因调控.它的命名与外星人ET无关,而是结合了expression和T7system.T7是溶菌型噬菌体.它在感染大肠杆菌後,首先利用宿主RNA聚合脢及核醣体制造出包含T7RNA聚合脢及T7蛋白激脢(proteinkinase)等噬菌体早期蛋白.T7蛋白激脢将宿主RNA聚合脢磷酸化使之失去活性;T7RNA聚合脢马上接管宿主细胞中所有的转录工作.T7RNA聚合脢是一条多胜链(singlepolypeptide)构成,只认识T7启动子不认得宿主启动子.载体pETs是将T7特有的启动子切出插入适当的载体中,利用这个启动子来启动欲表达的基因.至於T7RNA聚合脢则是选殖入乳醣分解脢基因启动子的後方,以λ噬菌体带入适当的宿主细胞内.当我们在培养基中加入IPTG诱发T7RNA聚合脢的大量制造时,大量的T7RNA聚合脢就转而大量地转录我们欲表达的基因.利用这种方式可以大量生产选殖在T7启动子後方的基因产物.生产量竟然能达到大肠杆菌蛋白质总量的1/10呢!这种载体称为表达载体(expressionvectors)
在生理学,遗传学及研究经验上,大肠杆菌及它的质粒和噬菌体都是我们最熟悉的.分子生物学及基因选殖也是从大肠杆菌开始发迹.基於这些原因,无论我们选殖动,植物基因或其他原核基因时,所考虑的第一个中间宿主总是大肠杆菌.在我们已经完成大肠杆菌中的基因选殖,鉴定及定序等工作後,选殖的基因总要送回原来的生物细胞或生物体中才有意义.然而绝大多数的大肠杆菌载体是无法在其它生物细胞中生存的.为了克服这种困难,於是人们纷纷切下包括动,植物及其它各种原核生物的质粒复制原,病毒复制原,染色体复制原,甚至染色体的部分DNA,将之插入大肠杆菌载体中.再配上其它适当的启动子,如此构筑成的载体不但可以用来转形大肠杆菌以外的细胞并且可以在这些细胞中做适当的基因表达.这一类能穿梭於两种不同生物细胞之间的载体便称为穿梭载体(shuttlevectors).
九,载体的适当利用
载体的种类繁多,厂商能够提供的及研究室或研究个人能够提供不计其数.当你需要利用载体时,你必须清楚你的目的.载体与宿主的搭配也非常重要,必须了解它们的特性与基因标示才能得心应手的运用.最後,也提供一些常用的质粒,载体与宿主来源:
机构:
ATCC: http://www.atcc.orgCABRI: http://www.cabri.orgCGSC/Yale University:http://cgsc.biology.yale.eduDSMZ: http://www.dsmz.deNational Institute of Genetics/Japan:http://shigen.lab.nig.ac.jp食工所生物资源保存及研究中心:http://www.ccrc.firdi.org.tw/ch-home.htm厂商:
Clontech: http://www.clontech.comFermentas: http://www.fermentas.comNew England Biolabs: http://www.neb.comNovagen: http://www.novagen.comPromega: http://www.promega.comRoche: http://www.roche-applied-science.comStratagen: http://www.stratagene.com