世界人种基因图谱31121 次点击159 个回复莉莉的脚 于 2012/6/17 18:16:29 发布在 凯迪社区 > 猫眼看人
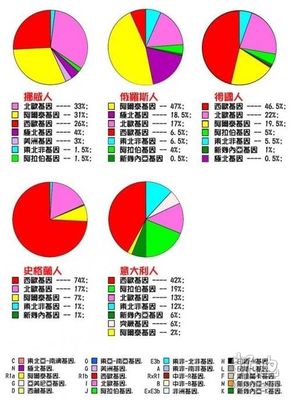
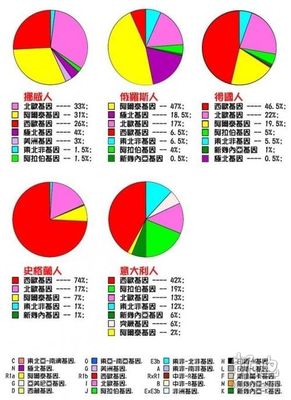
美国版人种基因图谱
图表来源:水木清华、凤凰网
先对表中的基因作一个说明:
黄种基因:N(极北基因)、O(东亚-南亚基因)
棕种基因:C(东北亚-澳洲基因)、D(矮黑人基因)
白种基因:F(斯里兰卡基因)、G(亚美尼亚基因)、H(印度-斯里兰卡H基因)、I(北欧基因)、J(阿拉伯-犹太基因)、K(新几内亚K基因)、L(印度-斯里兰卡L基因)、M(新几内亚M基因)、R1a(阿尔泰/印欧东支基因)、R1b(西欧/印欧西支基因)
黑人基因:A(东非-南非基因)、RxR1(中非R基因)、B(中非B基因)、ExE3b(非洲基因)
科普特-柏柏尔基因:E3b(东非-北非基因)
印第安基因:Q(美洲基因)
注意:图中白色标记的"突厥基因"是笔误,应改为"亚美尼亚基因"。
图表No.1
俄罗斯的阿拉伯基因来源于犹太人,俄罗斯现有200多万犹太人,此外还有大约200万有犹太血统的人,他们在沙皇俄国和苏联迫害时期被迫改变民族身份(改变信仰、族籍)。
俄罗斯人受东方印欧人的影响甚于伊朗人。
德国人有相当一部分是斯拉夫人后裔,如建立德意志帝国的普鲁士人。
意大利人的阿拉伯基因主要集中于南意大利,尤其是被阿拉伯人统治过300多年的西西里。
这份由美国人研究出的世界人种基因图谱(Y染色体)表明:现代德国人中属于雅利安基因占19.5%。而俄罗斯人属雅利安人基因占47%。雅利安人成份最高的是波兰,超过50%。以雅利安人自居的伊朗,雅利安基因只占18%。不如中国维吾尔族21%。印度操印欧语民族中雅利安基因占39.5%。
图表No.2
最不可思议:哈萨克人和蒙古人的大部分来源于澳洲原始白人。
蒙古有2%的伊斯兰教徒(基本上都是移居外蒙的回族)。
北方汉人和西藏人没有阿拉伯血统,研究人员可能对除了维族之外的其他穆斯林民族(比如回族、撒拉族)的历史缺乏了解,就凭蒙古人种的长相将这些民族归入到汉人和藏族的行列中了。目前中国有2000多万穆斯林,但据民间的说法,中国穆斯林人口大约在4000万至7000万之间(除去维族血统,阿拉伯血统差不多占北方人口的2%)。
西藏的阿尔泰基因来源于氐族(小月氏)。
维吾尔人的13.5%是驻守边疆的汉兵后代和吐蕃时期入侵新疆的汉藏语系部落后裔。
无论是从语言,考古还是DNA分析,汉人与现代的羌族都是很接近的。还有一点,那就是汉民族在黑暗时期的关于昆仑山昆仑神的传说恰好应证这一点。从某种意义上讲,汉人就是定居的羌人。之所以现代汉族对羌人没有认同感,理由同上。进入定居生活的华羌集团开始渺视那些还保持原始生活的羌人,慢慢在中国西边形成两大对立,生活方式截然不同的集团,即汉人和羌人。整个两汉王朝,汉羌不断兵戎相见。到南北朝,羌人一度在中国北方建立国家,称大秦。南北朝之后,羌人势力在中原衰微,转而向西南发展,形成西南众多民族。
图表No.3
土耳其人的极北基因可能是居住在西伯利亚的原始部落带来的。
亚美尼亚的阿拉伯后裔可能是拜占庭帝国安置的被迫改奉基督教的阿拉伯人。
中亚、西亚的黑人后裔有两种可能来源:一是被穆斯林统治者大量采购的黑人,二是伊拉姆人(埃兰人)的后代,但由于埃兰人已经消亡,前者很可能是大批来源。《一千零一夜》里就有许多黑奴伺候尊贵的阿拉伯主人。俄罗斯的黑奴在高加索还有后代。
中东阿拉伯人的东北非基因是原住民留下的。在阿拉伯帝国征服期间,中东有许多民族归依了伊斯兰教(如阿拉米人、巴比伦人、科普特人)。
印度印欧人来源于印欧语系东支。
11%的印度人信奉伊斯兰教。印度东北蒙古人种原住民中的米佐拉姆人被确认为犹太人后裔,全国2%的阿拉伯基因,他们可能能分到一份。此外阿富汗的杜兰尼人也被普遍怀疑为古代以色列“失踪的十族”。
印度的黑人大多属于安达曼群岛的土著。
【从DNA看人种历史】
一、现代人类起源
分子生物学技术的发展,使对DNA分子的直接研究成为可能。起先,一些有具体功能的基因的多态性(即人与人有差异)得到了关注。研究得最多的是人类特异性免疫蛋白基因HLA。他的确有相当多的类型,可以观察到大量群体间的差异。但由于与重要的人体功能有关,所以它的变异受到相当程度的限制,无法与人群的分化完全同步。其实,《人类基因组计划书》的完成使人们发现,在人类冗长的DNA分子中,有具体表达功能的基因只有三万个左右,而大部分的区段都只是填充基因组或维持分子结构的无意序列。这些序列的变异往往不会影响正常的人体功能,所以可以自由的与人群分化同步。然而,不管有无功能,位于常染色体上的序列都会碰到混血的难题。人类基因组主要分成23段,每一段构成一种染色体,其中22种是常染色体。每种常染色体在每个人体内都有一对,其一来自母亲,另一来自父亲,但在继续传给后代时,父母双方的染色体会打断而后重新拼接,即重组。所以不同位置的突变之间毫无关系,其组合完全没有意义。
于是,人们自然开始关注没有重组现象的区段,其一是线粒体,其二是Y染色体非重组区NRY。线粒体是男女都有的,但不管男女都来自其母亲,男性的线粒体无法传给后代(线粒体位于细胞质里,男女交配授精过程中,精子细胞只有细胞核和卵细胞核融合,精子的细胞质包括线粒体被排除在卵细胞之外),所以人们的线粒体都来自其女祖先,自然不存在重组。NRY则只有男性体内存在,更无法重组。这种不重组的区段,最大的优点是其上的突变是紧密相关的,每一个突变都代表着整个区段的特性。先后发生的突变,也会有上下游关系。这样,不仅突变的谱系清晰,不同性质的突变,在研究时也可以相得益彰。相关联的突变构成的组合结构叫做单倍型。而每种单倍型的发生年代可从其相关的”时钟”标记计算得到。这样,只要民族群体有特征单倍型,其发生渊源和年代就可推断。
但是这种构成单倍型的突变,必须保证只出现一次而且不会变回去,不然各种单倍型就不能保证其发生的独特性(同一种单倍型如有不同的来源,便不能据以分析群体发生的关系),有一种突变”核苷酸多态SNP”就符合这种要求。各SNP构成的某种单倍型都是单一来历的,NRY上的SNP单倍型就是在世界各地各民族中体现出极丰富的特异性和多样性。
另一种突变标记”短序列重复STR”与SNP单倍型相粘连时,就成了天然的”时钟”标记。在一个固定的基因位置上,有一种特定的短序列重复了几遍,有如火车车厢。每次突变都会增加或减少一节,而这种增加或减少是随机的。所以,同一个祖先的不同个体的同一STR位置,可能有不同的重复数。因为突变的速度是可知的,所以我们只要寻找到最大的重复数差异,再加上其它一些参数,就可以推算出他们共同祖先的生活年代。
不同位置上的STR,也可以构成单倍型。当几个STR同时随机增减时,奇妙的效果产生了。从最古老的单倍型开始,向不同方向产生差异越来越大的单倍型。在特定群体中观察到的中央单倍型总是最古老的,这就象一点墨水在清水中静静的扩散,色迹的中心必然是墨滴的最初位置。而根据扩散的半径,也可以计算扩散持续的时间。由于STR的突变机率比SNP大得很多,所以在同一种SNP单倍型的个体中分析STR的网络关系,就可以计算SNP的发生年代。这样,我们就掌握了民族遗传分析的两大利器。
由于技术难度不同,最早研究的非重组区是线粒体。在分析了遍及全世界的几千个人的线粒体之后,1987年,RebeccaCann发现线粒体的系统发源于非洲,这一学说被称为“非洲夏娃说”,这项研究揭开了遗传学方法探索人类史前历史的序幕。从现代人的线粒体发现现代人类起源于十万年前的非洲,那么其它大陆上更早时期的古人类句必然是已灭绝的旁支。这与古人类学界原有的普遍观点不合,因此引发了很多争论。但是,对欧洲尼安德特人骨骸DNA的分析,证实了他们的确与现代人没有传承关系,两个学界的认识也渐统一。
但是,由于线粒体的特殊性质,它的突变过快而且长度太短。这使得它提供的信息量很有限,而且在群体间的分布格局较乱,这一切则由NRY来捕捉。NRY上不断有新的SNP,到2001年,斯坦福大学的PeterA.Underhill等在全世界人群中发现了218个SNP位点,构成了131种单倍型。这些单倍型构成的谱系与线立体谱系的共同之处,也是证明现代人十万年前发源于非洲。
首先通过与黑猩猩等人类近亲的相关区段的比较,我们找到了人类谱系的”根”的方向,因为SNP的特异性,人类的SNP位点,在黑猩猩的对应位置上必然未突变。所以与黑猩猩单倍型最接近的人类SNP单倍型,必然是人类最古老的形态。而后随着时间的推移,不断有新的人类的单倍型演化出来。结果发现所有最早演化出来的新态都保存在非洲,而非洲之外的人类的单倍型形态只是较晚(约12万年前)发生的一小支。这说明现代人最早(约15万年前)的演化过程发生在非洲,直到大约12万年前,才有一小部分人走出非洲,向全世界扩散。
这种结构在NRY的谱系中表现得比线立体清晰和稳定得多。由于NRY是纯粹父系遗传的,所以这种非洲起源假说被称为“非洲亚当说”。人们往往对非洲人的黝黑肤色难以认同,其实现代非洲人已不是世界上其它人种的直系亲属。只是在公元1000年左右,西非洲的黑人(Y染色体单倍群E)掌握了农业技术,人口增长而后扩张,才把东非洲等地的浅色的“我们的近亲(Y染色体单倍群A和B)”取代了。而由于欧洲人于大航海时代的殖民,西非洲的这种取代,就被打乱而未能完全完成,所以南非的土著肤色不太黑。
东亚的古人类学界至今还有人不相信。北京猿人竟然不是我们的祖先。但是在这里,的确没有找到104万年前的人类化石,看起来这里的古人种,在约10万年前就灭亡了。在之后的几万年冰河时期,东亚大地寒冷而寂寞,直到四万多年前,来自非洲的现代人重新发现了这块大地。
人的血型的一大特点是:终身不变,且能遗传。按照孟德尔遗传定律,父亲双方各遗传一种基因给子女,子女的血型里就包含着这些基因,因此,每个人的血型既受之于父母,但又不同于父母,通过验血型可以认定父母与子女间的血缘关系。而且,人类的这种血缘关系源远流长,早在人类祖先从猿人分化出来时,一种叫Gm血型的因子已形成,它代代相传,延续至今。可见,在血型里,不仅有父母的标记,而且还有老祖宗的标记。
人体血型主要有红细胞血型、白细胞血型、血小板血型、血清血型和红细胞酶型五大类,总共有500种以上。ABO血型已为大家所熟知。但ABO血型仅是红细胞的血型,而携带着人类种族特征的血型是血清的血型。血清血型要比红细胞血型复杂,它有几十种之多,其中,叫做Gm的血清血型(双倍体遗传的常染色体基因)有明显的种族差异。
Gm血型的种族差异,表现在组成Gm血型的各个因子上。组成Gm血型的各个因子,目前世界上已检出的是18个,其中有的因子是不同人种所专有的,如fb因子只存在于白种人,st因子为黄种人特有,C3因子仅见于黑种人等等;此外,在同一人种的不同民族中,Gm因子的基因频率具有特定值,即是占有某种Gm因子的人数在总人数中占有比较恒定的百分比。如我国的壮族,具有afb因子人的百分比总在80%左右,而维吾尔族人中是7%左右。由基因频率特定值可以确定民族间的血缘关系。Gm血型的这两大特点,是至今发现的人体500多种血型中独一无二的。这使它成为研究种族关系的重要依据。
人类的东非祖先的一支后裔Y染色体单倍群突变为A-M91。
12万年前,人类的东非祖先的另一支后裔Y染色体单倍群突变为B-M60和M168。
M168继续突变为三个分支:
C-M130、P9(YAP)、F-M89。
P9(YAP)又继续突变为D-M174和E-M96(SRY)。
10万年前,C和D(Gm血型是ag)最先走出非洲,形成棕色人种。他们沿着印度洋岸边,拿着旧石器和尖木叉,在海边捕鱼为生,并捡食滩涂上的高蛋白食物。经过了一代又一代人的扩展,来到了印度,形成了前达罗毗荼人ag/axg。前达罗毗荼人是所知的棕色人种中文化发展的最为先进的一支,与后来的L共同创造了哈拉巴文化
接着,棕色人种东继续东进,到了东南亚。当时的海平面比现在要低好多,巺他群岛与中南半岛连成一片,澳大利亚和伊利安岛以及美拉尼西亚的许多岛屿也连在一起。6万年前,棕色人种到达并占领了这片地区。到后来海平面抬升以后,澳大利亚和伊利安岛远离亚洲大陆,棕色人种在这两块孤立的大陆上居住到了现在。4万多年前,棕色人种又北上占领整个东亚大陆,创造了中国的旧石器时代晚期文化,如山顶洞人、资阳人。当时的日本列岛与朝鲜半岛、美洲与亚洲都是连在一起的,棕色人种沿着今对马海峡到达日本,成为绳文人,并且沿着今白令海峡到了美洲。
中国北方(包括西北、华北、东北、外蒙古、朝鲜)的棕色人种,与2万多年前之后从中亚零星迁入的吐火罗人『按:吐火罗最初源于语言学研究,吐火罗语这个名称对不对还是二说,中国学者倾向于叫它焉耆龟兹语,吐火罗人何时迁到新疆没有定论(但肯定是在印欧语形成之后),不知道本文作者是否在臆断,抑或此处的吐火罗是个人种学的概念,和说焉耆龟兹语的人不是同一个民族,个人猜测黄帝可能是焉耆龟兹人的同族,这种语言属于印欧语西支和拉丁语日耳曼语等有密切关系,虽然焉耆龟兹人地处今日中国境内,但和印欧语东支的伊朗印度语差别很大,英国语言学家w.亨宁认为他们是灭亡苏美尔阿卡德王朝德蛮族kutim人.逐渐融合形成了阿尔泰民族(Gm血型是吐火罗ab3st+棕色人种ag/axg),所以阿尔泰民族Y染色体特色单倍群以C为主,以P为辅。在东北、华北,吐火罗的因子特别少,形成通古斯群(比较纯种的棕种人,Y染色体单倍群基本是C);在蒙古高原东部,吐火罗与棕色人种在人口比例上平分秋色,形成蒙古群(Y染色体单倍群C和P各占一半);在新疆、蒙古高原西部、黄土高原,吐火罗的影响力比较深,形成突厥群(Y染色体单倍群是C和P)。
此时整个东亚、南亚大陆及澳洲只分布着棕种人的Y染色体单倍群C、D型。今天,C只在亚洲和美洲土著有发现,里海以西没有C的存在。D可能先于C来到东亚大陆,是东亚最早的原住民,后来到达的C对其进行了驱逐与覆盖。所以D只分布在边缘的岛屿和高寒的西藏。
二、白种人
7万年前,黄白种人F-M89(P14)(Gm血型是ab1b3)也离开非洲,他们没有走海岸线,而是走陆路,6万年前到达西亚(苏美尔、伊朗、巴基斯坦)并定居在那里,形成欧亚人群(Gm血型突变为afb1b3)。目前,F占德拉维达(达罗毗图)诸民族的13%、印度印欧语系的3%、伊朗族的5%、乌孜别克族的7%、哈萨克族的4%、喀山鞑靼的4%、吉尔吉斯族的2%、阿尔泰族的2%、藏族的1%、蒙古族的1%。
接着,G-M201、H-M69(M52)、J-M304(1212)从欧亚人群分离出去,G-M201迁移到意大利、小亚细亚、伊朗,今天的意大利族的6%、伊朗族的4%、土耳其族的7%拥有G;H-M69(M52)迁移到印度,今天操印欧语系的印度人的20%、德拉维达(达罗毗图)诸民族的18%拥有H;J-M304(1212)是阿拉伯的祖先,随着阿拉伯人的扩张,今天中东阿拉伯族的48%、埃塞俄比亚族的15%、达罗毗图诸民族的12%、维吾尔族的10%、哈萨克-吉尔吉斯族的5%、乌兹别克族的8%、蒙古族的2%、回族的8%、俄罗斯族的3%、土耳其族的25%、伊朗族的12%、意大利族的20%、墨洛哥的15%、苏丹的10%拥有J。
I-P19(M170)也从欧亚人群分离出去,迁移到北冰洋沿岸,今天冰岛族的37%、苏格兰族的20%、挪威族的30%、拉普人的30%、德意志族的25%、西班牙族的12%、西班牙族的15%、意大利族的15%、伊朗族的14%、中东阿拉伯族的5%、俄罗斯族的18%、土耳其族的15%、喀山鞑靼的15%、涅涅茨人的6%、鄂温克族的6%、科里亚克族的7%拥有I。
K-M9也从欧亚人群分离出去,由于东部高山的影响,K-M9分为两个方向继续向东迁徙,一部分L-M20(M11)向南,进入印度次大陆。今天,印度操印欧语系的人26%、操德拉维达(达罗毗图)语系的30%、维吾尔族的3%、乌孜别克族的2%携带L,中东人也有1-2%携带L。3万年前L与从南部沿海进入印度的“海上移民”棕色人种C4、C5遭遇,其间发生了激烈的冲突,新来的侵略者L与棕色人种的女人们交配,而那些棕色人种的男人们则大部分遭到杀害。今天C4、C5占整个印度(包括印欧语系和达罗毗图语系)诸民族的比例为5%,但在个别达罗毗图语系民族中比例达到20%。今天的德拉维达(达罗毗图)诸民族就是L、H与C、F的融合。
K-M9另外一部分M-M5(M4/H17)继续东进,来到澳洲,今天伊里安查亚的70%、巴布亚新几内亚的34%拥有M。K-M9的另外一部分K则进入澳洲和中国,今天苏门答腊岛的15%、婆罗洲的16%、伊里安查亚的5%、巴布亚新几内亚的59%、澳洲土著的25%、汉族的10%拥有K。
K-M9另外一部分P-P27(M45)于3.5万年前向北面的兴都库什山走去,继续向北进入中亚里海北岸游牧,即今天的印欧语系。印欧语系西支染色体突变为R-M207(M173),Gm血型由afb1b3突变为fb1b3。公元前3000年,印欧语系西支R1b离开故乡分好几路向四面八方扩张。西支一路到了希腊,西支二路到了意大利(这两路分别是古希腊和古罗马文化的起源)。西支三路穿过中欧一直到达不列颠诸岛(凯尔特人的祖先)。西支还有两路北上到了东欧波罗的海,西边的为日尔曼、东边的为斯拉夫。这些西支白种人每到一处,就征服或同化当地的土著(尼安德特人),把自己的语言传播到那里。公元前1000年,最后一拨印欧语系西支R1a(M17/H16)也离开里海北岸的故乡,一路南下分别占领了伊朗、苏美尔、阿富汗和达罗毗图语系的印度,他们都号称雅利安人。这拨人里面还有一部分(RxR1)从印度出发,继续往西南行进,远抵喀麦隆。
今天,冰岛族的40%、苏格兰族的75%、德意志族的48%、西班牙族的65%、意大利族的44%、挪威族的25%、俄罗斯族的6%、土耳其族的14%、涅涅茨人的4%、塞尔库普人的4%、拉普人的6%、中东阿拉伯的5%、伊朗族的5%、喀山鞑靼的4%、乌孜别克族的7%、维吾尔族的17%、回族的12%携带R1b。
今天,俄罗斯族的47%、挪威族的35%、冰岛族的22%、德意志族的22%、拉普人的20%、印度讲印欧语的40%、讲德拉维达(达罗毗图)语的8%、阿尔泰族的50%、喀山鞑靼的18%、乌孜别克族的18%、维吾尔族的18%、哈萨克族的5%、吉尔吉斯族的40%、涅涅茨人的10%、塞尔库普人的18%、拉普人的20%、伊朗族的13%、土耳其族的6%、中东阿拉伯的2%、藏族人的5%、回族的5%携带R1a。
今天,德拉维达(达罗毗图)语的8%、印度操印欧语的3%、维吾尔族的4%、乌孜别克族的4%、吉尔吉斯族的2%、哈萨克族的1%、土耳其族的1%、伊朗族的1%、澳洲土著的6%、喀麦隆土著的45%拥有RxR1。
印欧语系东支染色体没突变,仍然是P,但Gm血型由afb1b3突变为ab3st。今天,P主要分布在中亚,其次印度的分布比例也较高(与中亚人历次军事南侵和商旅往来有关),最后中东与东亚也有零星分布(主要是当时中东和东亚人口众多,而当时的印度人口不算太多)。印欧语系东支于2万年前向东迁移,到达新疆及蒙古高原,称吐火罗人,他们后来全部同化于突厥人、蒙古人之中,但却给其带来了P的染色体和ab3st的Gm血型,并使突厥人由棕种-黄种人变成为黄棕白混血人种。
西伯利亚最初是土著棕色人种C,后来I又移民过来,再后来阿尔泰人(P、C)又移民过来,最后黄种人N也移民过来,最终融合形成雅库特人(N为主)、楚科奇人、科里亚克(N、C为主)人、尼夫赫人、尤卡吉人、勘察加人等古西伯利亚人(北亚人)。1.5万年以前,部分北亚人(母系是黄种人但父系却是白种人P)又经过白令陆桥进入了美洲,成为阿留申人、爱斯基摩人(N占50%,Q3占47%)、印地安人(基本上是Q3)。北亚人在向西伯利亚、美洲行进过程中,一部分到达美洲,Y染色体从P突变为Q3-M3(H15);一部分从西伯利亚南下中原,Y染色体从P突变为Q1-M120,混入中原通古斯,再后来成为东夷最西一部、最后成为今日河南汉族的一部分。
三、黄种人
5万年前,K-M9中的L-M20(M11)、P-P27(M45)白人分离出去之后,剩余的人群染色体突变为M214,接着又分为N、O,但Gm血型仍然是afb1b3,而没有象印欧人一样突变为fb1b3。N、O沿喜马拉雅山南麓匆匆通过,进入东南亚,这期间也融入了少量棕色人种的基因。剩余的N、O的另一支留居西亚(苏美尔、伊朗、巴基斯坦)故土,到后来创造了人类最早的文明苏美尔文明、克里特岛文明和伊朗-阿富汗的先雅利安文明。
4万年前,在东南亚的密林里,N、O演变为黄种人。黄种人逐渐发展壮大了起来,在与棕色人种的对比中,渐渐占据了优势,并最终将自己居住区域(华南-东南亚)的棕色人种不分男女全部予以种族灭绝,而不是同化融合。也就是说,此时华南南部和东南亚随着黄种人的到来,Y染色体发生了C、D被O的替换。
2.5—3万年前,黄种人并分化成了沿海支、内陆支、乌拉尔支。内陆支黄种人又称南亚(百濮)/澳斯特罗-亚细亚,形成于缅甸一带,并散布于整个中南半岛(除占城)、中国云南省、广西省和海南岛,今中国的佤-德昂-布朗、缅甸-泰国的孟族、柬埔寨的高棉族、越南的京族、马六甲族、尼科巴族、印度的蒙达族、斯里兰卡的维达人都是其后代。由于历史比沿海支稍长,渐渐形成了更多的基因突变,南亚(百濮)的Y染色体特色单倍群为O2(包括M88/O2a1+M111)和O3(包括M7/O3d+M134/O3e)。百越民族中,泰国泰族的65%、中国壮族的35.95%也是O2的后代。
沿海支黄种人又称百越(侗台)南岛,Y染色体单倍群是O1-M119(包括M50+M110/O1b+M103)。沿海支黄种人最初在越南占城一带形成,其中的百越民族离开占城北迁至广东(如柳江人),其中的南岛民族离开占城南迁至马来半岛。在广东,百越民族居住了几万年而没有外迁,因此发育出了统一的体质和文化。百越分成南北两部分,南部是壮傣和黎的祖先,北部是侗水和仡央的祖先。
当时,第四纪晚更新世冰期(距今7万年)不仅仅造成了海平面下降、大陆边缘延伸,另一个效应是一些不高的山都是积雪冰封,使得南岭等山脉成了黄种人向北进入长江流域的天然障碍。1万年前,冰期结束,使得黄种人大量北上,开创了一个全新的新石器时代,农业文明的星星之火在各地的黄种人中燃烧了起来,社会发展的火车头被拉动了。黄种人的北上分两条路线进行。
西部内陆支的百濮民族通过贵州、四川这条路线北上,并且分出了两支。一支沿着长江往下走,到了长江中游(以江汉平原为中心,南包洞庭湖平原、西尽三峡川东、北达豫南豫中、东抵鄱阳湖的广大地区),形成南蛮(苗瑶),Y染色体是O2a*,后来汉藏部落O3d混入南蛮,而南蛮的O2a1则脱离南蛮混入南越。南蛮先后创造了石门皂市下层文化(为前5000)→大溪文化(前4400前3300)→屈家岭(湖北京山县)文化(前3000—前2000)→湖北龙山文化(青龙泉三期,前2400,与中原龙山文化有较大区别)四种新石器文化。百濮另一支沿着四川的几条大河(金沙江-雅砻江-大渡河)北上,(另一种途径折返向西经喜马拉雅山南麓达阿富汗,然后经新疆天山、昆仑山、祁连山东进)来到了黄河上游地区,形成了汉藏民族,Y染色体是O3,特别是O3e(M134)。
由于广西、云南、中南半岛、海南居住着百濮,湖南北部居住着苗瑶,所以东部的沿海支的百越民族最早的扩散方向是空旷无人的东北和正东。北部百越(以M119亚类型M110为特色)向东北迁徒进人江西,形成干越。干越一部溯长江而上向西远迁到贵州西北部,成仡央族群,在商朝时建立了夜郎国。在商末,楚国发动了对江西干越的征伐,干越大部分人退到广西梧州,被称为骆越。秦汉时候,部分骆越迁到了广西北部的融水一带,又渐向贵州移动成为今天的水族。宋代受越南侵略军的侵扰,骆越又从梧州北上广西、湖南、贵州三省交界地带,成为今天的侗族。部分干越后来东迁进人浙江,形成了於越,在太湖平原和宁绍平原创造了灿烂的新石器文化。於越先后创造了浙江余姚河姆渡文化(前5000~前4000年)、太湖流域马家浜文化(分为马家浜和崧泽两期,为前4300~前3300年)、良渚文化(前3300~前2200年,与山东龙山文化有更密切的联系)、南京北阴阳营文化、安徽潜山薛家岗第三期文化(前3200~前3000年)。于越民族继而又从江苏进入山东,与当地的苗瑶、通古斯融合创造了先东夷文化。
南部百越(混入了部分苗瑶的O2a1-M88)部分也在北部百越离开后一段时间开始内开始东进,到达福建和浙南,形成后来的闽越和东瓯。西汉南越灭国后,汉朝时武帝灭南越,南越开始大规模西迁,到唐朝时进入西双版纳、老挝、泰国。1215年更从云南的勐卯国(现云南德宏)西迁至印度阿萨姆省建立了阿含Ahom王国。南越小规模进入海南岛也较早,但是大规模进入海南是较晚的事,形成黎族。台湾原住民是不同时期沿不同路线从大陆来到台湾的,少部分属于南越,大部分属于北越。今壮族的17.9%、泰国泰族的10%、越南京族的7%、柬埔寨高棉族的8%、土家族的20%拥有M119。
同属黄种人东部沿海支的南岛民族,与同胞百越民族相反,他们不是向北,而是向南面广阔的大洋进发。公元前3000年之后,逐渐从占城、马来半岛扩散到菲律宾、印尼、马来西亚。公元后又进一步扩散到马达加斯加、密克罗尼西亚[马绍尔、吉尔伯特(基里巴斯)、关岛查莫罗、加罗林群岛(波纳佩-雅普-特鲁克)、帕拉乌、瑙鲁]、美拉尼西亚[斐济、莫图、雅比姆、所罗门群岛、瓦努阿图]、波里尼西亚[新西兰毛利、乌维亚、萨摩亚、汤加、图瓦卢(艾利斯)、钮埃、库克群岛拉罗通加、社会群岛塔西提、土阿莫图、马克萨斯、夏威夷]。
四、黄种人进入北方取代棕色人种
1万年前,沿海支、内陆支的黄种人分别北上,阿尔泰在与吐火罗人(Y染色体单倍群为P)第一次混血的基础上,又与当时北上的两支黄种人发生了第二次混血,形成黄棕混血的黄种人北亚人种。东部阿尔泰通古斯混有较多的沿海支成分O1,但也混有很多内陆支成分O3(可能是进入文明时代后加入的);中部阿尔泰蒙古混有较多的内陆支成分O3;西部阿尔泰突厥也开始混入了大量的内陆支成分O3。
目前,蒙古族血统中,C3占55%,O3占20.5%,O1占4.2%,O2占1%,K占4%,N占5%,R1a、D、Q、J各占2%,RxR1占1%,F占0.8%,R1b占0.5%。朝鲜族血统中,O2占32%,O3占25%,O1+O*占14%,C1占16%,D占7%,N占2%。满族的Y染色体类型中,C1占23%、C3占2%、D占6%、N占15%、O占53%。维吾尔族有10%的阿拉伯血统J、3%的印度黄白血统L、39%的雅利安血统R、4%的蒙古血统C、4%的藏族矮黑血统D、2%的芬兰黄种人血统N、14%的汉朝唐朝戍守西域的汉族士兵血统O3。
C1-M8占鄂温克族的65%、鄂伦春族的61%、日本大和族的8.5%、阿伊努人的90%、朝鲜族的15%、满族的25%(含C3)、赫哲族的30%。C3-M217占布里亚特蒙古族的60%、蒙古族的55%、达斡尔族的31%、锡伯族的27%、哈萨克族的45%、乌孜别克族的17%、吉尔吉斯族的10%、维吾尔族的4%、阿尔泰族的23%、雅库特族的11%、土耳其族的1%、喀山鞑靼的1%、古西伯利亚科里亚克族的30%、回族的23%、藏族的2%、印度讲印欧语的3%、塞尔库普的3%。C2—M38占澳大利亚土著的70%、伊里安查亚的25%、巴布亚新几内亚的3%、婆罗洲的17%。C4、C5占印度(包括印欧和达罗毗图语系)的比例为5%。
D分布于印度安达曼群岛土著(100%)、苏门答腊岛土著、冲绳人(55%)、藏族(50%)、土家族、彝族、缅甸族、克钦/景颇族、瑶族、日本(34.7%)、朝鲜(7%)、满族人(6%)、维吾尔族(4%)、阿尔泰族(4%)、蒙古族(2%)、乌孜别克族(2%)、哈萨克族(1%)。N-M231所占比例中,布里亚特为27%、叶尼塞鄂温克族17%、中国鄂温克族0%、鄂伦春族的5%、赫哲族18%、满族15%、达斡尔族9%、锡伯族21%、蒙古族5%、阿尔泰族4%、哈萨克族7%、吉尔吉斯族3%、喀山鞑靼21%、乌孜别克族2%、维吾尔族2%、日本2%。
O1—M119所占比例中,布里亚特蒙古人5%、锡伯族7%、达斡尔族4%、蒙古族4.2%、满族5.6%、乌尔奇Ulchi(赫哲亲属)3.8%、雅库特4%、回族的8%、古西伯利亚尼夫赫6%、堪察加人的6%、科里亚克人7%、日本人3.4%、叶尼塞鄂温克3.2%、中国鄂温克族5%、朝鲜族的14%(含O*)。
O2—M95主要分布在泰国-柬埔寨-越南南部的南亚(百濮)语系和中国的苗瑶语族;而且,百越民族如泰国泰族的65%、中国壮族的35.95%也是O2;另外,东北亚O2含量也很高,如日本的O2比例高达31.7%,朝鲜族O2比例亦高达32%,满族的O2比例为11%,赫哲族为5%、达斡尔族为21%、鄂伦春族5%、叶尼塞鄂温克0%、中国鄂温克族2%、锡伯族1%、回族的2%、蒙古族1%。
O3所占比例中,蒙古族20.5%、锡伯族30%(含O*)、达斡尔族27%(含O*)、满族3**%(含O*)、赫哲族46%(含O*)、鄂伦春族22%(含O*)、朝鲜族25%、日本人20.1%、维吾尔族14%、哈萨克族8%、吉尔吉斯族24%、回族17%(含O*),中国鄂温克族28%(含O*)、叶尼塞鄂温克族和布里亚特蒙古人为0。
阿尔泰人在黄土高原(突厥)的故土被内陆支的黄种人(汉藏民族)霸占,生活在那里的阿尔泰人不是被屠戮就是向北面的家园逃窜,所以今天黄土高原的居民很少有阿尔泰特有单倍群C的发现。
阿尔泰人在华北(通古斯)的故土遭到了沿海支黄种人的大规模入侵,不过,沿海支黄种人相对比较温和,与通古斯和平共处而不是屠戮,二者逐渐融合为新的民族-东夷,东夷的O1成分浓厚。
新石器时代,阿尔泰人在阴山以北创造了以细石器/刮削器在生产工具中占优势的狩猎文化,与阴山以南以农耕文化为主的仰韶文化有明显不同。商族的母系祖先东夷从燕山地区南下到古河济之间的进程中,一部分阿尔泰人(蒙古群)也随之从大兴安岭山原南下来到商族祖居的赤峰市、朝阳市、承德市、北京市,创造了拉苏克游牧文化之一的夏家店上层文化。
后来的匈奴、坚昆、丁零、金山突厥等突厥部落,都是黄种人居于统治地位的棕色白色混血人种。随着突厥人的西迁中亚,他们与当地的晚于吐火罗人到来的印欧语系的雅利安人也发生融合。公元400年突厥族群的匈奴西迁,演化为保加尔人。公元600年蒙古族群的柔然人厌达人西迁,一部分被斯拉夫人同化,一部分被保加尔人同化。保加尔人的一支演化为目前的楚瓦什人,保加尔人的另一支被斯拉夫人同化为保加利亚人。中亚北部的突厥族群可萨人、佩切涅格人、乌古思人、钦察人逐渐演化成目前巴什基尔人、库梅克人、诺盖人;后来一部分与西迁的蒙古人融合为鞑靼人。
黄种人乌拉尔支N-M231(M178)跟随内陆支黄种人来到黄河上游,并进入突厥地盘,与O3一起征服了突厥,成了那里的主宰,原始突厥语、汉藏语、乌拉尔语混杂在一起,形成了一种新的语言-乌拉尔语。前3000年,N-M231途经阿尔泰山,最终西迁至乌拉尔山,形成乌拉尔族群。乌拉尔族群与当地的占人口多数的斯拉夫-日尔曼人(R)、波罗的海人(I)融合,形成芬兰-乌戈尔人,再后来分成三支向外扩散:向北的一支沿着伏尔加河上游进入北欧,演变为芬兰人、芬兰以北的拉普人(N占45%,I占30%,R1a占20%,R1b占5%)、爱沙尼亚人、莫尔多维亚(莫尔多瓦)人、乌德穆尔特人、马里人、卡累利阿人、科米人(黄种人);向西的一支演变为马扎尔人;向东一支,沿卡马河、越过乌拉尔山,进入西伯利亚,演变为汉蒂(奥斯恰克)人、曼西(沃古尔)人等。剩下的乌拉尔族群被称为萨莫耶德人,包括涅涅茨人(分布在北西伯利亚和乌拉尔山脉两侧,N占80%,R1a占10%,I占6%,R1b占4%)、塞尔库普人(N占7%,R1a占18%,R1b占6%,C占3%,Q占66%)、牙纳桑人,他们生活在俄罗斯极北地区,是相对纯粹的黄种人。
黄种人沿海支和内陆支大规模来到北方以后,分别对当地的阿尔泰-棕种男人进行了杀戮和驱逐,但保留了阿尔泰-棕种女性(Gm血型是ag+axg+ab3st)以婚配,所以在一定程度上可以说,北迁的黄种人的后代与阿尔泰-棕种的后代是同母异父。当历史进入了阶级社会,情况就发生了变化,奴隶的需求使得黄种人对遭遇到的棕种男人不再灭杀。那段时期黄种人才到达的边缘地区,比如青藏高原、蒙古高原、日本列岛,棕色人种的成分被大量地保留了下来。
日本列岛在棕色人种绳文人到来时还可以通过大陆架很容易地到达,而当黄色人种来到东北亚时,已成为茫茫大洋相隔的海岛,是黄种人传说中的蓬莱仙岛。现在北海道的虾夷人(阿伊努人)和种子岛的部分居民就是绳文人的后代。日本的黄种人弥生人,是春秋战国之后从中国大陆迁入的东夷人(O2+O1)、汉族人(O3)、通古斯人(C1),弥生人在九州岛登陆,然后向本州岛及更东北方向扩张的过程中,对旧石器的绳文人进行了征服和奴役,造成了日本绳文人比例的下降。但是到1800年,本州岛的北部和库页岛,都还是虾夷人的领地。从日本各地的遗传结构看,棕色人种的遗传比例虽然是从西往东递减,但各地的该种成分都不在少数。所以日本人总体上应该算是两个人种的混血。日本大和民族的Y染色体类型中,O-M175占54.2%(其中O2M95占31.7%,O3M122占20.1%,O1M119占3.4%);D占34.7%;C1-M8占8.5%;N占1.6%。阿伊努人的Y染色体类型中,C1-M8占90%,D占10%
五、东夷的华夏化
原始东夷属于苗瑶、百越、通古斯民族的混血,特有的是Y染色体单倍群是O2、O1、C1,分布在华北平原东部和山东半岛辽东半岛,北方的东夷有更多的通古斯血统,南方的东夷有更多的苗瑶、百越血统,但总体上以苗瑶、百越为主。自从东夷首领与西邻原始华夏族在中原地区接触以来,东夷混入了越来越多的原始华夏的因子,特别是华北平原的东夷已经大幅度华夏化(不但指语言上而且指血统上)。
东夷以凤为祖图腾,繁衍为以各种鸟为子图腾的诸多部落。原始东夷创造了先进的海岱文化:发明了带羽毛弓箭、创造了东夷文字(对汉字的诞生有启蒙作用)、制作青铜器与冶铁、制造舟-车、发展农业与治水。为紧邻民族华夏之文化的发展和推进,起到了关键和决定性作用,中原华夏文明就是文化相对落后的西部华夏族吸收先进的东部东夷族文化后进入文明社会进而在周朝超越东夷文化的。从伏羲、神农、炎黄直到夏商二代,在华夏-东夷文化(主要指科技,不包括语言)互动的发展中起主导作用的也是东夷文化,但在政治上语言上起主导作用的还是华夏族(商与东夷联合灭夏,受东夷影响很大)。
太昊(即伏羲氏后裔分支)姓风,是原始华夏首领伏羲氏派驻东夷的殖民代表,东夷化成了东夷的世袭首领。春秋时的任(济宁市)、宿-须句(东平县)、颛臾(费县西北)都是太昊后裔回到伏羲母系故里后建立的方国。神农氏部落与太昊部落(该族分两支,原支在淮阳发展为华夏族,东迁一支在曲阜发展为东夷)一样,也分为淮阳和山东两大系统,淮阳以华夏语言为主、山东以东夷语言为主,但神农氏各部落后来全部为东迁的炎帝部落(四岳之一支)所并。炎帝四岳之一支东迁到今鲁西豫东接壤地区,以淮阳为都,完全取代了原神农氏的地盘,与太昊集团交错分布;居住在山东的四岳本为华夏,后来全部东夷化(语言上)。太昊集团有华夏族龙崇拜,但同时也和东夷一样以鸟为图腾崇拜。
蚩尤就是东夷的子孙,但具体是太昊还是神农之后,不可考。
少昊是东夷继太昊之后的又一个统治集团,嬴姓。少昊不是一个人,而是一个集团。帝喾的儿子少昊挚只是东夷诸多少昊首领中的一个,并且与之前和之后的少昊首领也没有父系的血缘关系,母系倒有可能。少昊挚是华夏族派驻东夷的殖民代表,后来东夷化(改姓东夷姓嬴)变成东夷的代表了。
嬴姓祖少昊,其后裔春秋时仍有徐、江、葛、黄、淮夷、费、郯、谭、锺离等国。徐,今安徽泗县和江苏泗洪;江,今河南正阳县西南;黄,今河南潢川县西北;葛,今河南宁陵县;费,今山东费县;郯,今山东郯城县;谭,今山东章丘县境;锺离,今安徽风阳县。少昊嬴姓的一部分西迁在晋陕立国,成为秦国、赵国的祖先。偃姓祖皋陶,与少昊同姓,嬴、偃同字异写,由于部落的分化所致。偃姓在西周春秋时尚有奄(曲阜县),为少昊所都;英,今安徽金寨县;六,今安徽六县;舒鸠,今安徽舒城县;东夷群舒,均为偃姓。莒国(今山东莒县)先为嬴姓,后改己姓,是少昊集团加入黄帝集团的部落。
夏初,启与伯益争夺共主地位的斗争,中经后羿、寒浞代夏政,直到后杼灭有穷氏,前后经历半个世纪的斗争,是少昊集团中那些与炎黄集团结成部落联盟的各部落在部落联盟向国家过渡过程中的斗争。这些部落已从东夷中分化出来,加入了华夏雏形形成的行列,成为华夏起源时期东系的重要来源。他们分布在古河济之间,即今鲁、豫、冀三省接壤的地区。在夏代,还有一支从东夷中分化出来加入了华夏雏形形成行列的,就是商族(父系是华夏,母系是东夷)。他们从燕山地区南下,兴起于河济之间,终于造成了代夏而建立中国第二个王朝的基础。在泰山与古济水流域,形成了以奄(今曲阜)与薄姑(今山东博兴县)为中心的各方国。西周初奄与薄姑仍是东夷中的大国,他们是东夷中社会发展水平最接近夏、商的方国,而泰山以东至海,以南至淮的各部落与方国,即夏商时期的九夷或夷方。
西周初既灭奄及薄姑,封鲁与齐于其故地。以齐鲁为中心,今山东省境内到春秋的只有莱夷没有完全华夏化。淮水地区诸夷,为西周东方的劲敌,是西周中后叶东夷的主体。齐与鲁对周围东夷部落的政策有明显的区别:齐太公治齐“因其俗,简其礼”,鲁公伯禽治鲁“变其俗,革其礼”。齐所采取的政策,促进了华夏文化与东夷文化的融合,不仅在春秋时首先称霸,齐国的文化对整个华夏文化的影响也不可低估。齐国周围的东夷部落较快地与之融为一体,前567年齐国灭莱,于是山东半岛各东夷小国已被齐兼并。鲁国伯禽强调变革当地土著居民的礼俗,虽然鲁以推行周礼着称,但当地土著东夷的习俗仍顽强地保留着。鲁在东夷文化起源发展的中心地区,以华夏族之周文化为主导,发展出对后世有很大影响的孔孟儒家学说。孔夫子,其先为宋人,是商族苗裔。
今江苏北部、安徽北部分布着少昊集团的后裔嬴姓与偃姓诸国,如徐、江、葛、黄、淮夷、锺离、英、六、舒鸠等,他们是蚩尤战败后由山东迁移过来的东夷部落,但南迁后含有更多的百越血统。春秋时期他们与诸夏通婚,参与会盟,在战国大兼并的历史进程中,他们先后被楚、鲁、吴-越所兼并,他们的文化对齐、鲁、楚及吴越都有影响,在政治上逐步统一过程中,文化与民族得到交汇融合,夷夏间的差别逐渐消失,成为华夏族的一部分。
应该说一直到汉朝,东夷的华夏化才彻底完成,东夷在向华夏族完全贡献了自己的科技智能后,在血缘上、语言上被华夏族函化,最终成为汉族的一分子。所以目前山东、苏北、皖北之汉族,血缘上都含有通古斯和百越血统,非纯粹的华夏血统,但人种上与华夏无异,属于北方远东人种。
原始汉人创造前仰韶文化时,东夷人创造了北辛(山东滕县)-青莲岗(江苏淮安市)文化,分布于淮北、鲁南,为前5400~前4400。
原始汉人创造仰韶文化时,东夷人创造了大汶口文化(是北辛-青莲岗文化的继续),主要分布在全山东、江苏淮河以北,在皖北、豫东和辽东半岛也有零星分布,为前4300~前2500年。大汶口文化的早期受到仰韶文化的影响。到了晚期,它还深入豫中地区,如平顶山寺冈、偃师滑城等地。辽东半岛旅顺郭家村下层,是大汶口文化向北波及的证据。
华夏族创造中原龙山文化时,东夷人创造了山东龙山文化(典型龙山文化,是大汶口文化的继续)。山东龙山文化的分布以山东为主,也包括江苏、河北和辽东半岛的部分地区,为前2500~前2000年。
华夏族创造夏-先商-先周文化时,东夷人创造了岳石(山东平度县东)文化(前1900—前1500)。岳石文化填补了山东龙山文化与商文化的空隙。
东夷被华夏族击败后,东夷北支逃到辽河流域,深深影响了西伯利亚和北亚居民的基因构成,O1—M119成为阿尔泰语系民族中普遍存在的基因,O2成为满族、朝鲜族、大和族的第一基因;东夷南支向南逃窜,回到自己的百越老家,并将部分C1/D带到南方。从此,东夷民族从肉体上、文化上从黄河流域消失,取代他的是带有O3—M122的华夏族。
六、华夏族的形成
汉藏民族Y染色体单倍群是O3—M122。O3出现频率最高地区为中国云南省、印度东北部那加邦的藏缅族群里,很多民族中出现频率为100%,如独龙族等。在靠近中国的东南亚地区,O3数量比较多,缅甸、老挝、越南都超过50%。
汉藏民族一支形成了藏缅族群,他们创造了马家窑文化,以陇西为中心,西至河西走廊和青海省北部,北达宁夏南部,南抵四川省北部。马家窑文化是陕西仰韶文化的一种地方性的变体,年代为前3300前2050年。齐家文化起源于中原之陕西龙山文化,随着不断向西发展,分布与马家窑文化地域重合,开始承袭了马家窑文化的若干因素,地域性也就显得更加突出,实属陕西龙山文化的变体。齐家文化早期为前2000年,已普遍出现铜器,晚期年限可到商代。至于甘肃西部的火烧沟文化,则已进入比较发达的青铜时代,为前1900~前1700。
藏缅族群不断南下,占领了原属棕色人种的和兄弟民族百濮的很多地盘,并于之通婚,融合成现在的藏族人、羌族人、氐族人、彝族人、缅甸族人、蜀人、巴人(土家族)人。
其中,藏族是秦汉时期藏缅族群中的两支唐旄发羌(仙女),南下进入青藏高原,与当地的棕色人种土著雅砻(猕猴)混血而形成的。现在的藏族人的遗传结构还能看到大量的D型Y染色体(当然O3型染色体占50%以上)。而观察藏族的面貌特征,甚至在同一个村子中,也会看到两种截然不同的样式,有的人是挺拔的羌人的面容,有的人则是敦圆的棕色人种面貌,肤色的差异也很大。
氐族现在仅存于四川和甘肃边界地区,有着惊人的Y染色体遗传结构全部的D型,说明他们是棕色土著的后代,并且在汉藏语中,“氐”就是“底层”的意思。羌人来到青藏高原,把原本就在这里的棕色土著称为“氐族”,因为他们“原抵”就在那里。羌族也含有部分的D型,但还是以O3为主。另外,整个藏缅族群由于在黄河上游老家跟C融合的厉害或者因为后来匈奴、鲜卑、金山突厥、契丹、蒙古的影响,所以现在都含有10%以上的C的血统。
O3在南亚语群体和苗瑶语群体中就存在。唯有O3e1是汉族才独有的,这个类型诞生于原始汉人的故乡黄土高原(甘南宁夏陕北),很可能是做了800年江山的周族的祖先帝喾-后稷(弃)。今日北方汉族O占73%,K占13%,C、Q各占4%,R1a、J、其他各占2%;今日南方汉族O占71%,K占14.4%,C占8%,D占4%,F占1%,R1a占0.8%,N占0.8%。